

A Two-Step Process of Effector Programming Governs CD4+ T Cell Fate Determination Induced by Antigenic Activation in the Steady State
- PMID: 33238127
- PMCID: PMC7714042
- DOI: 10.1016/j.celrep.2020.108424
A Two-Step Process of Effector Programming Governs CD4+ T Cell Fate Determination Induced by Antigenic Activation in the Steady State
Abstract
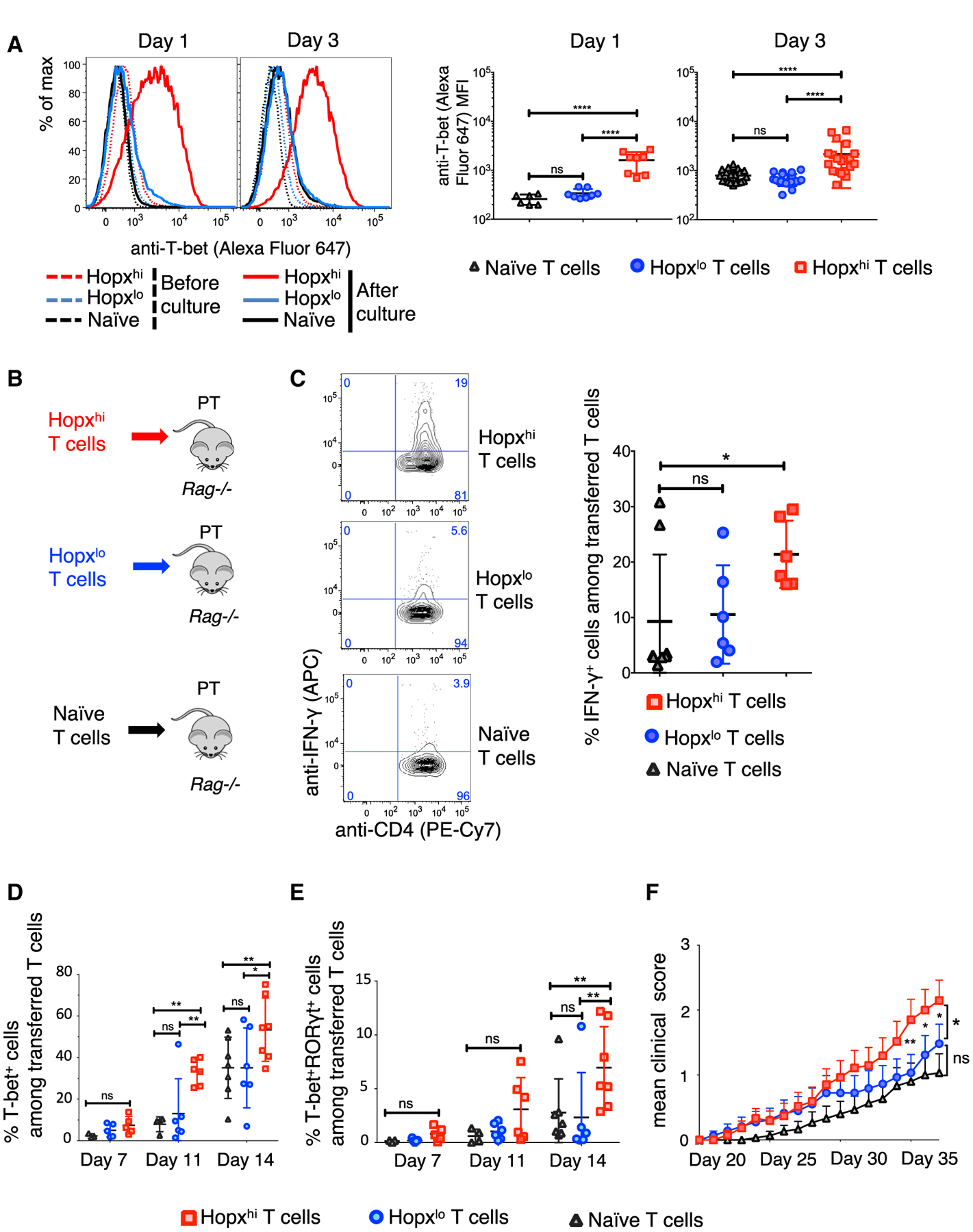
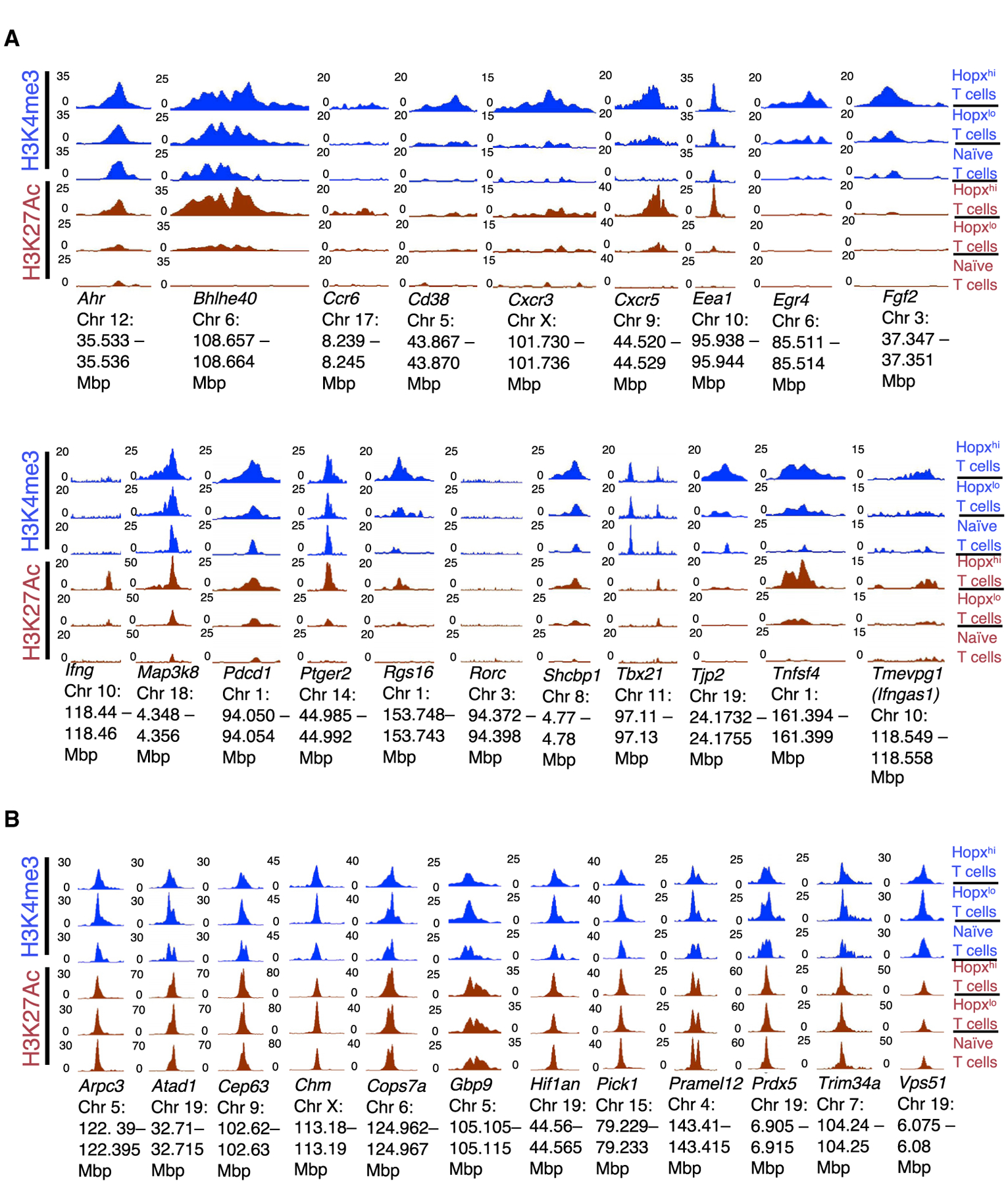
Various processes induce and maintain immune tolerance, but effector T cells still arise under minimal perturbations of homeostasis through unclear mechanisms. We report that, contrary to the model postulating primarily tolerogenic mechanisms initiated under homeostatic conditions, effector programming is an integral part of T cell fate determination induced by antigenic activation in the steady state. This effector programming depends on a two-step process starting with induction of effector precursors that express Hopx and are imprinted with multiple instructions for their subsequent terminal effector differentiation. Such molecular circuits advancing specific terminal effector differentiation upon re-stimulation include programmed expression of interferon-γ, whose production then promotes expression of T-bet in the precursors. We further show that effector programming coincides with regulatory conversion among T cells sharing the same antigen specificity. However, conventional type 2 dendritic cells (cDC2) and T cell functions of mammalian target of rapamycin complex 1 (mTORC1) increase effector precursor induction while decreasing the proportion of T cells that can become peripheral Foxp3+ regulatory T (pTreg) cells.
Keywords: EAE; Hopx; IFN-γ; T cells; autoimmune; dendritic cells; effector programming; mTORC1; steady state; tolerance.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures



References
-
- Albrecht I, Niesner U, Janke M, Menning A, Loddenkemper C, Kühl AA, Lepenies I, Lexberg MH, Westendorf K, Hradilkova K, et al. (2010). Persistence of effector memory Th1 cells is regulated by Hopx. Eur. J. Immunol 40, 2993–3006. - PubMed
-
- Ardouin L, Luche H, Chelbi R, Carpentier S, Shawket A, Montanana Sanchis F, Santa Maria C, Grenot P, Alexandre Y, Grégoire C, et al. (2016). Broad and Largely Concordant Molecular Changes Characterize Tolerogenic and Immunogenic Dendritic Cell Maturation in Thymus and Periphery. Immunity 45, 305–318. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
