

Transformative Network Modeling of Multi-omics Data Reveals Detailed Circuits, Key Regulators, and Potential Therapeutics for Alzheimer's Disease
- PMID: 33238137
- PMCID: PMC7855384
- DOI: 10.1016/j.neuron.2020.11.002
Transformative Network Modeling of Multi-omics Data Reveals Detailed Circuits, Key Regulators, and Potential Therapeutics for Alzheimer's Disease
Abstract
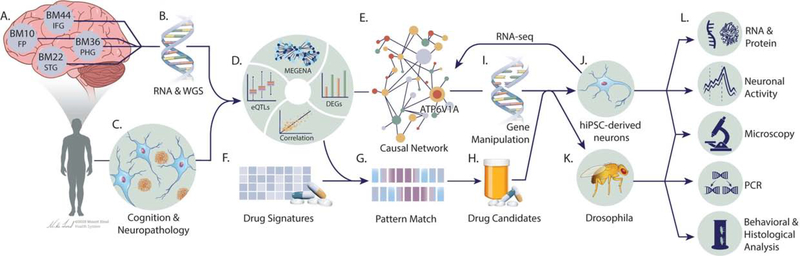
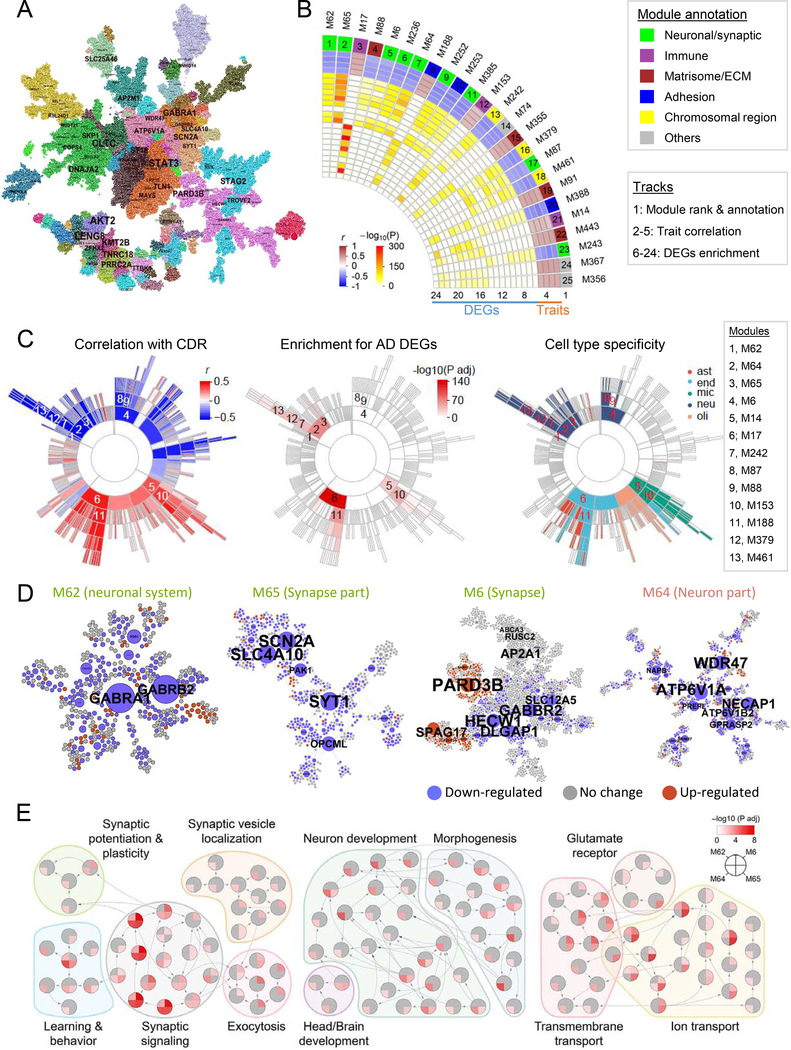
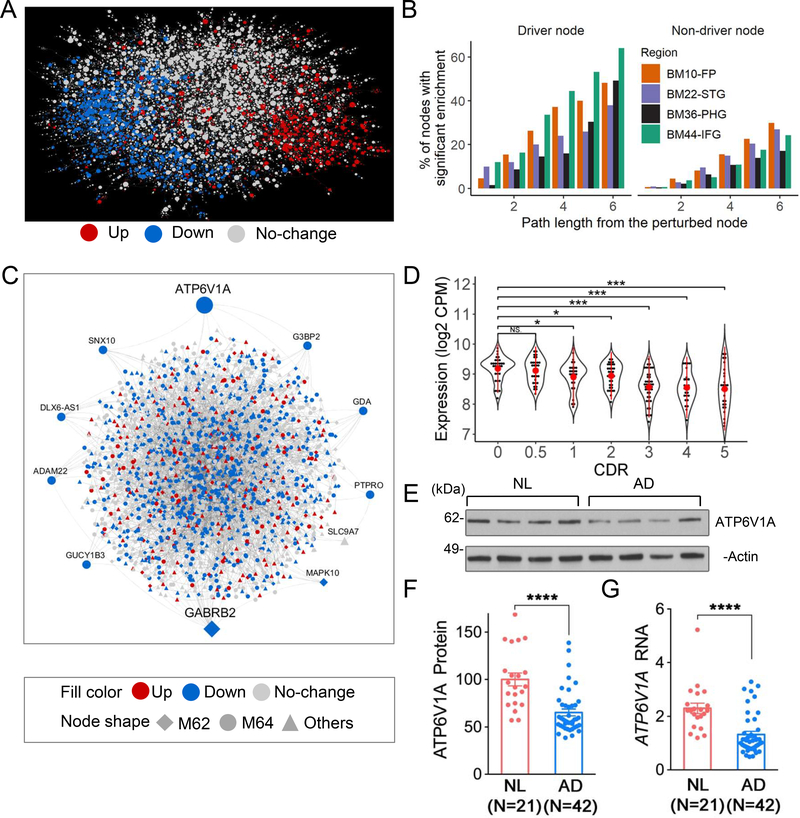
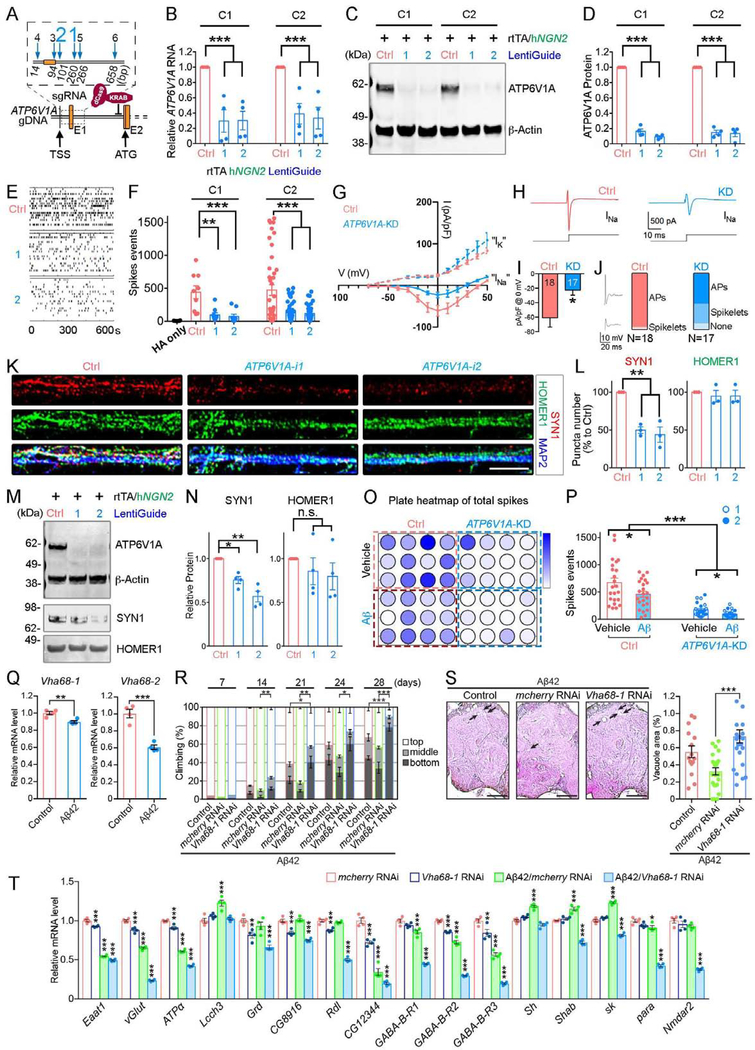
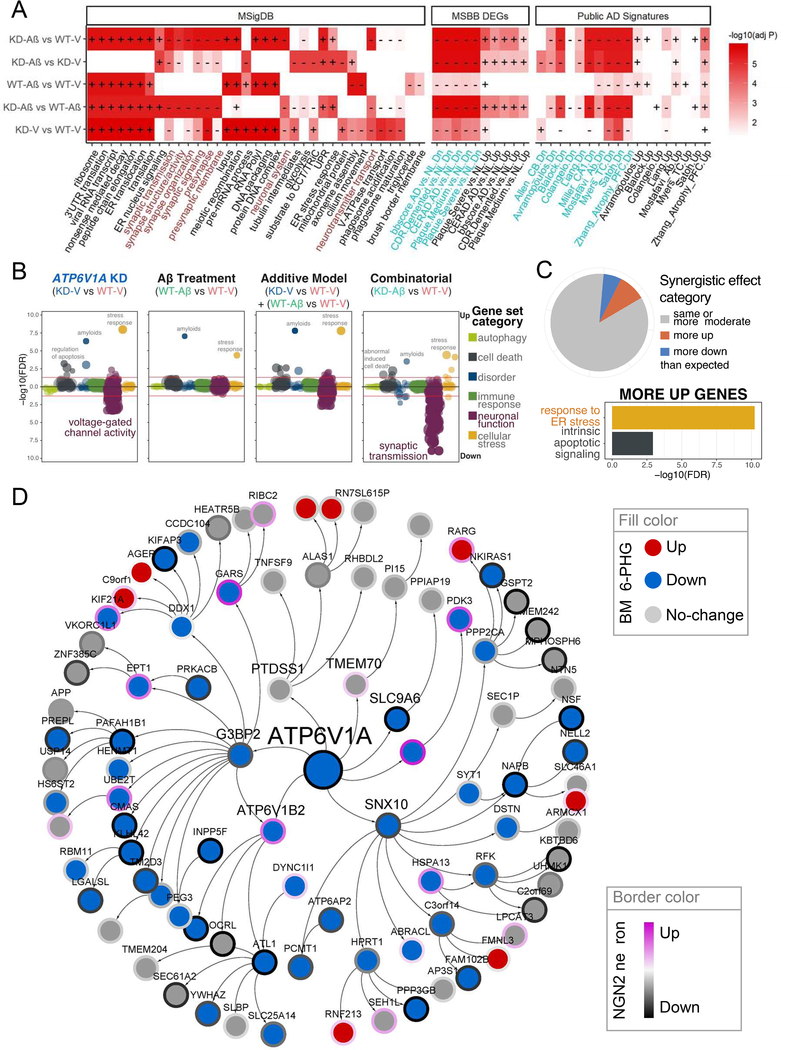
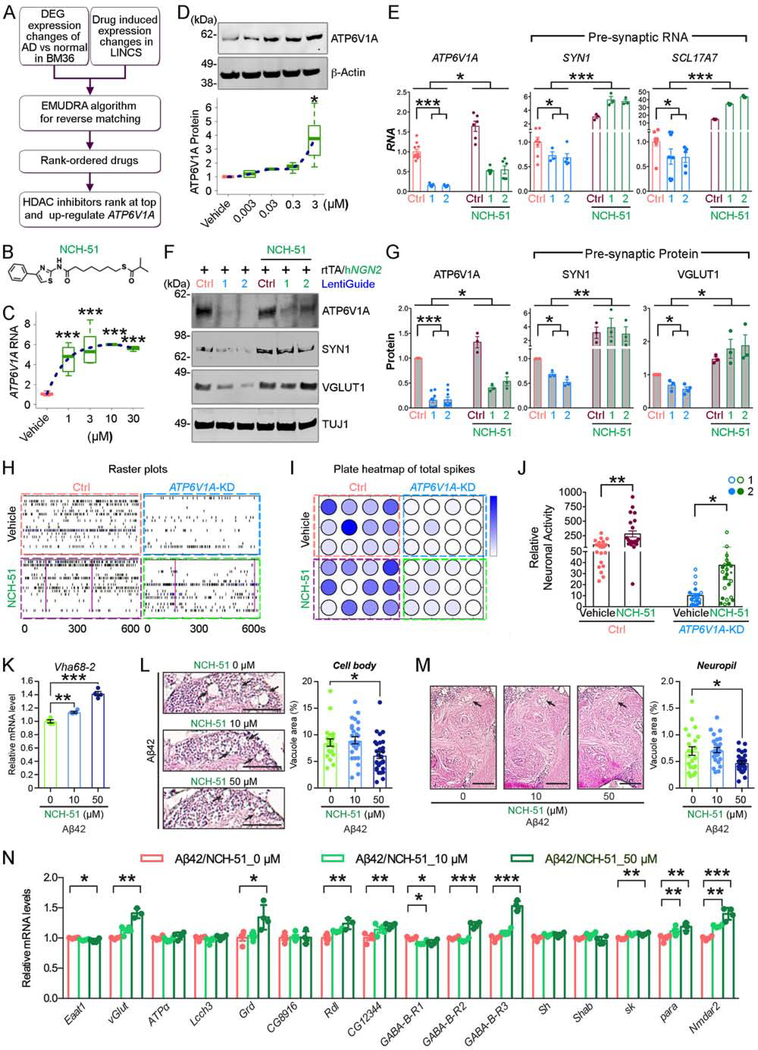
To identify the molecular mechanisms and novel therapeutic targets of late-onset Alzheimer's Disease (LOAD), we performed an integrative network analysis of multi-omics profiling of four cortical areas across 364 donors with varying cognitive and neuropathological phenotypes. Our analyses revealed thousands of molecular changes and uncovered neuronal gene subnetworks as the most dysregulated in LOAD. ATP6V1A was identified as a key regulator of a top-ranked neuronal subnetwork, and its role in disease-related processes was evaluated through CRISPR-based manipulation in human induced pluripotent stem cell-derived neurons and RNAi-based knockdown in Drosophila models. Neuronal impairment and neurodegeneration caused by ATP6V1A deficit were improved by a repositioned compound, NCH-51. This study provides not only a global landscape but also detailed signaling circuits of complex molecular interactions in key brain regions affected by LOAD, and the resulting network models will serve as a blueprint for developing next-generation therapeutic agents against LOAD.
Keywords: ATP6V1A; Alzheimer’s disease; Drosophila; NCH-51; NGN2 neurons; human induced pluripotent stem cell; network biology; neuronal dysregulation; omics.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
Comment in
-
An Integrated Multi-omics Approach Identifies Therapeutic Potential for ATP6V1A in Late Onset Alzheimer's Disease.Neuron. 2021 Jan 20;109(2):193-194. doi: 10.1016/j.neuron.2020.12.015. Neuron. 2021. PMID: 33476559
References
-
- Ando K, Maruko-Otake A, Ohtake Y, Hayashishita M, Sekiya M, and Iijima KM (2016). Stabilization of Microtubule-Unbound Tau via Tau Phosphorylation at Ser262/356 by Par-1/MARK Contributes to Augmentation of AD-Related Phosphorylation and Aβ42-Induced Tau Toxicity. PLoS Genet 12, e1005917. - PMC - PubMed
-
- Association A.s. (2018). 2018 Alzheimer’s disease facts and figures. Alzheimer’s & Dementia 14, 367–429.
Publication types
MeSH terms
Grants and funding
- R01 AG072562/AG/NIA NIH HHS/United States
- RF1 AG057440/AG/NIA NIH HHS/United States
- RF1 AG054014/AG/NIA NIH HHS/United States
- U01 AG052411/AG/NIA NIH HHS/United States
- R01 AG068030/AG/NIA NIH HHS/United States
- R56 AG058655/AG/NIA NIH HHS/United States
- U01 AI111598/AI/NIAID NIH HHS/United States
- RF1 AG057443/AG/NIA NIH HHS/United States
- RF1 AG044546/AG/NIA NIH HHS/United States
- P30 AG066514/AG/NIA NIH HHS/United States
- R01 AG062355/AG/NIA NIH HHS/United States
- R56 MH101454/MH/NIMH NIH HHS/United States
- U01 AG046170/AG/NIA NIH HHS/United States
- R01 AG057907/AG/NIA NIH HHS/United States
- I01 BX003380/BX/BLRD VA/United States
- RF1 AG058501/AG/NIA NIH HHS/United States
- I01 RX002290/RX/RRD VA/United States
- R01 DA043247/DA/NIDA NIH HHS/United States
- RF1 AG059319/AG/NIA NIH HHS/United States
- U01 AG058635/AG/NIA NIH HHS/United States
- R01 DK118243/DK/NIDDK NIH HHS/United States
- R03 DE026814/DE/NIDCR NIH HHS/United States
- R01 MH121074/MH/NIMH NIH HHS/United States
- RF1 AG048923/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
