

NLR-1/CASPR Anchors F-Actin to Promote Gap Junction Formation
- PMID: 33238150
- PMCID: PMC7725993
- DOI: 10.1016/j.devcel.2020.10.020
NLR-1/CASPR Anchors F-Actin to Promote Gap Junction Formation
Abstract
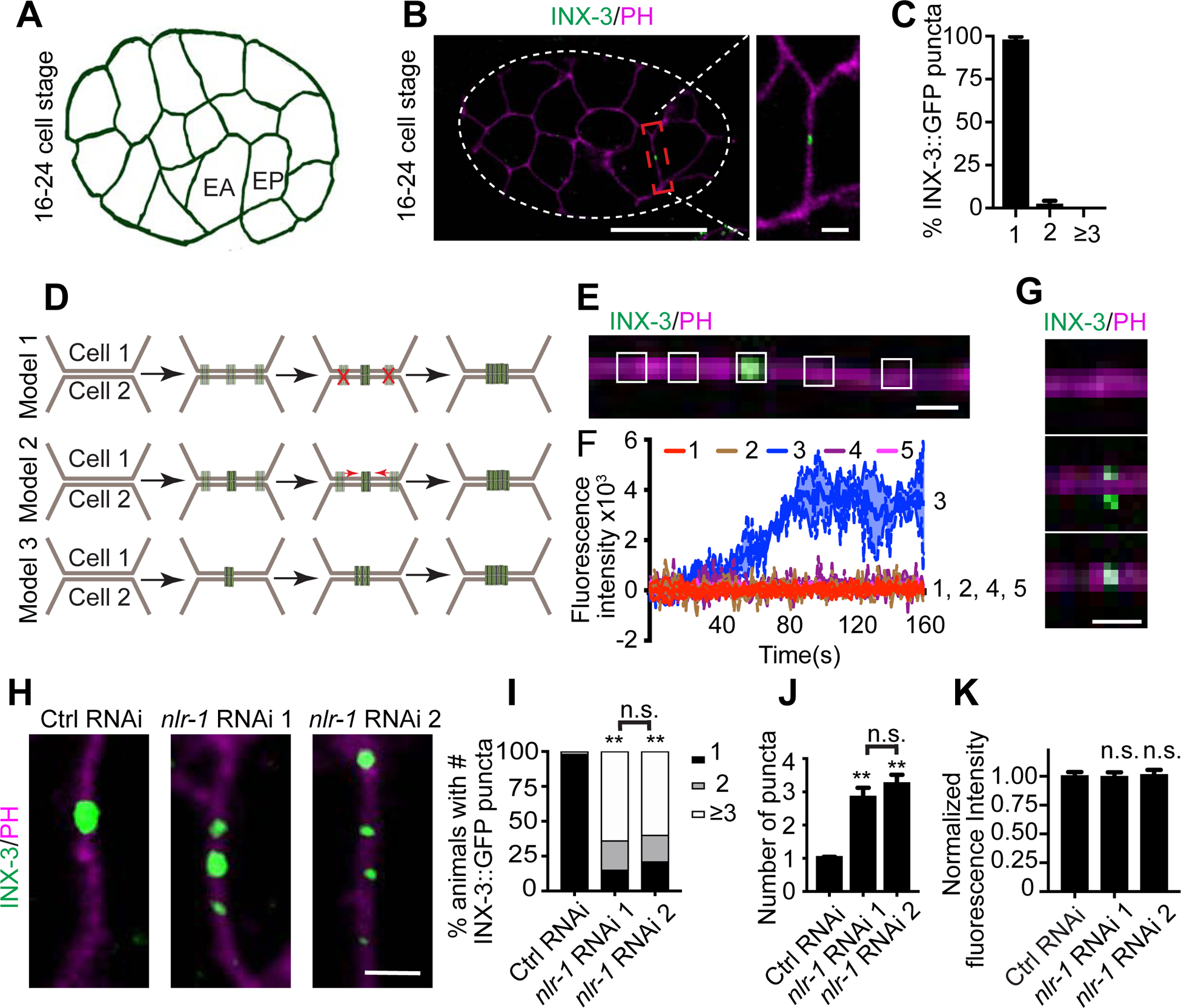
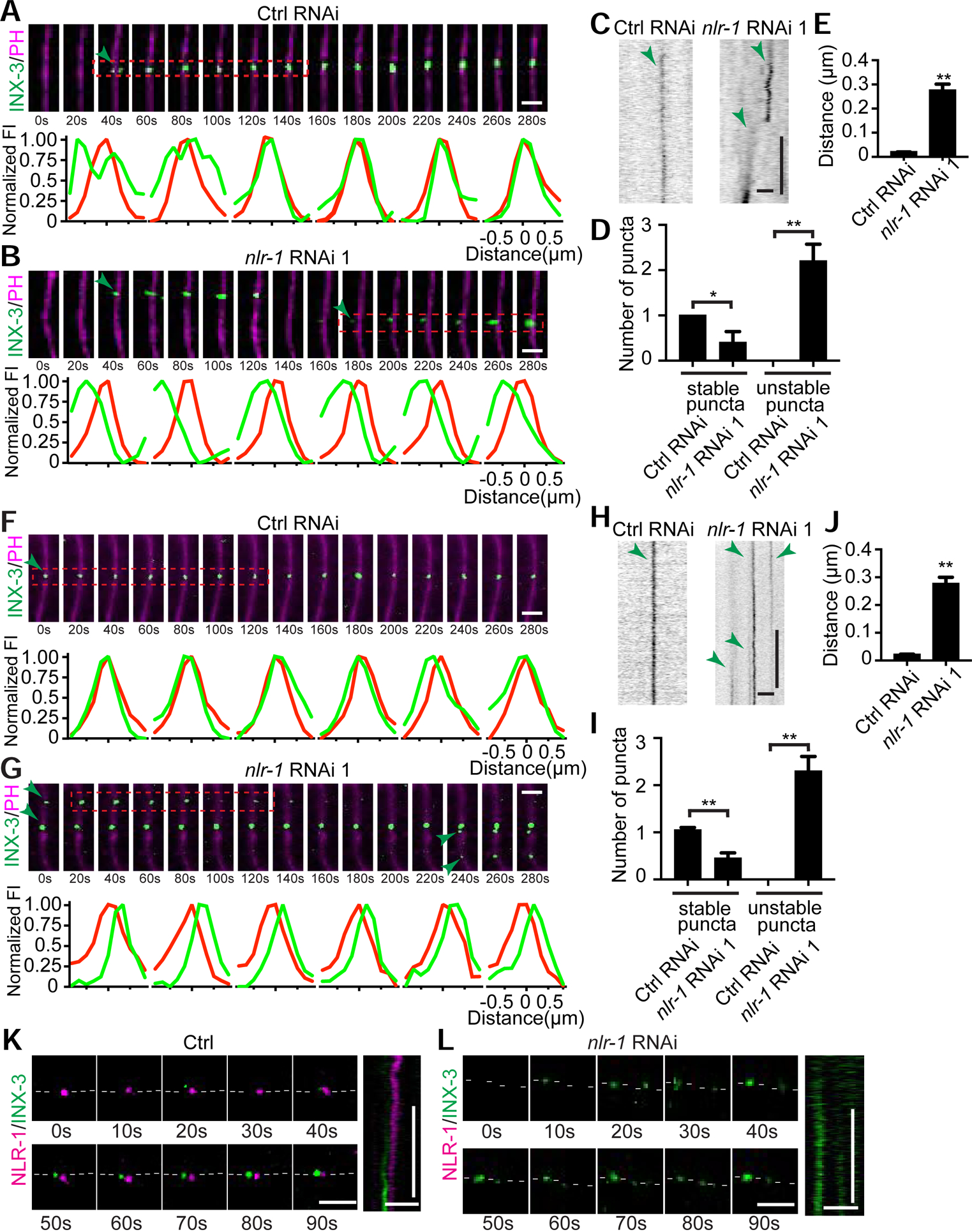
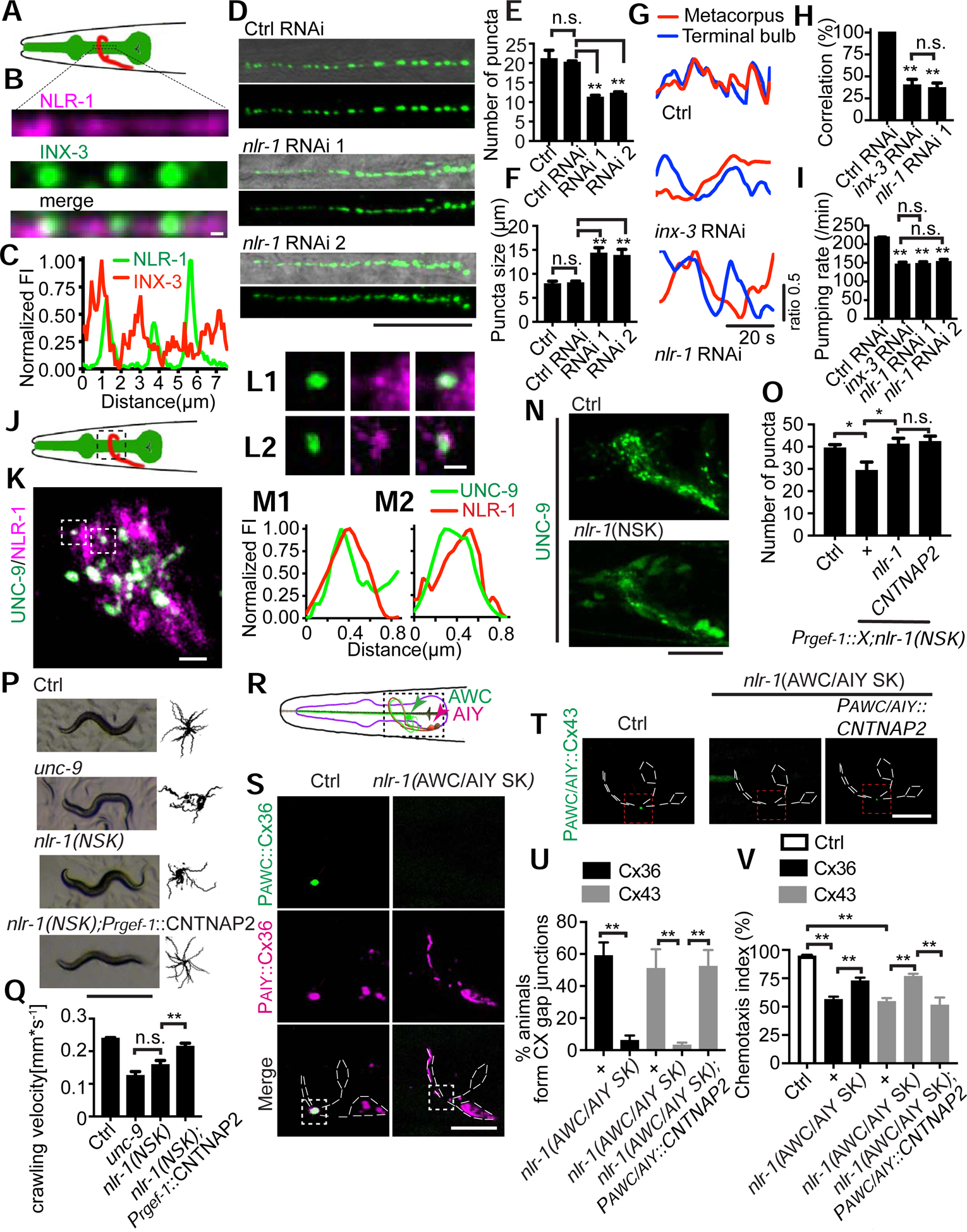
Gap junctions are present in most tissues and play essential roles in various biological processes. However, we know surprisingly little about the molecular mechanisms underlying gap junction formation. Here, we uncover the essential role of a conserved EGF- and laminin-G-domain-containing protein nlr-1/CASPR in the regulation of gap junction formation in multiple tissues across different developmental stages in C. elegans. NLR-1 is located in the gap junction perinexus, a region adjacent to but not overlapping with gap junctions, and forms puncta before the clusters of gap junction channels appear on the membrane. We show that NLR-1 can directly bind to actin to recruit F-actin networks at the gap junction formation plaque, and the formation of F-actin patches plays a critical role in the assembly of gap junction channels. Our findings demonstrate that nlr-1/CASPR acts as an early stage signal for gap junction formation through anchoring of F-actin networks.
Keywords: C. elegans; F-actin; Gap junction; contactin-associated protein.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures


References
-
- Albertson DG, and Thomson JN (1976). The pharynx of Caenorhabditis elegans. Philos Trans R Soc Lond B Biol Sci 275, 299–325. - PubMed
-
- Alexopoulos H, Bottger A, Fischer S, Levin A, Wolf A, Fujisawa T, Hayakawa S, Gojobori T, Davies JA, David CN, et al. (2004). Evolution of gap junctions: the missing link? Current biology : CB 14, R879–880. - PubMed
-
- Bauer R, Loer B, Ostrowski K, Martini J, Weimbs A, Lechner H, and Hoch M (2005). Intercellular communication: the Drosophila innexin multiprotein family of gap junction proteins. Chemistry & biology 12, 515–526. - PubMed
-
- Bauer R, Weimbs A, Lechner H, and Hoch M (2006). DE-cadherin, a core component of the adherens junction complex modifies subcellular localization of the Drosophila gap junction protein innexin2. Cell communication & adhesion 13, 103–114. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
