

The cochlear ear horn: geometric origin of tonotopic variations in auditory signal processing
- PMID: 33239701
- PMCID: PMC7689495
- DOI: 10.1038/s41598-020-77042-w
The cochlear ear horn: geometric origin of tonotopic variations in auditory signal processing
Abstract
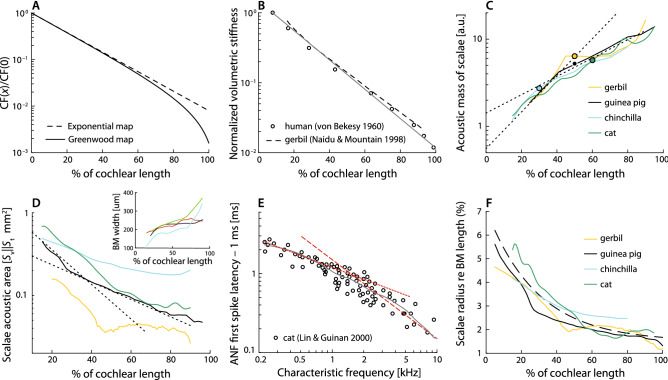
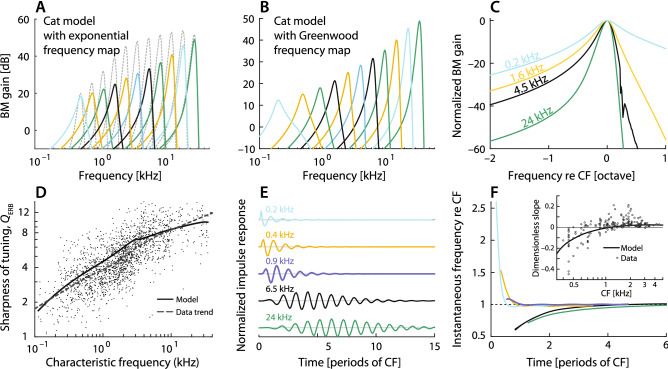
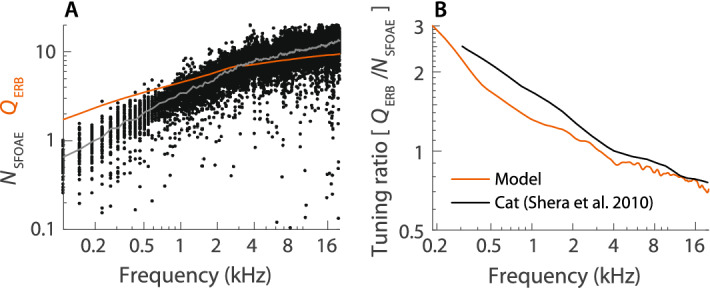
While separating sounds into frequency components and subsequently converting them into patterns of neural firing, the mammalian cochlea processes signal components in ways that depend strongly on frequency. Indeed, both the temporal structure of the response to transient stimuli and the sharpness of frequency tuning differ dramatically between the apical and basal (i.e., the low- and high-frequency) regions of the cochlea. Although the mechanisms that give rise to these pronounced differences remain incompletely understood, they are generally attributed to tonotopic variations in the constituent hair cells or cytoarchitecture of the organ of Corti. As counterpoint to this view, we present a general acoustic treatment of the horn-like geometry of the cochlea, accompanied by a simple 3-D model to elucidate the theoretical predictions. We show that the main apical/basal functional differences can be accounted for by the known spatial gradients of cochlear dimensions, without the need to invoke mechanical specializations of the sensory tissue. Furthermore, our analysis demonstrates that through its functional resemblance to an ear horn (aka ear trumpet), the geometry of the cochlear duct manifests tapering symmetry, a felicitous design principle that may have evolved not only to aid the analysis of natural sounds but to enhance the sensitivity of hearing.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Rhode WS, Cooper NP. Nonlinear mechanics in the apical turn of the chinchilla cochlea in vivo. Aud. Neurosci. 1996;3:101–121.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
