

An autosomal dominant neurological disorder caused by de novo variants in FAR1 resulting in uncontrolled synthesis of ether lipids
- PMID: 33239752
- PMCID: PMC8026396
- DOI: 10.1038/s41436-020-01027-3
An autosomal dominant neurological disorder caused by de novo variants in FAR1 resulting in uncontrolled synthesis of ether lipids
Erratum in
-
Correction to: An autosomal dominant neurological disorder caused by de novo variants in FAR1 resulting in uncontrolled synthesis of ether lipids.Genet Med. 2021 Dec;23(12):2467. doi: 10.1038/s41436-021-01189-8. Genet Med. 2021. PMID: 34667295 Free PMC article. No abstract available.
Abstract
Purpose: In this study we investigate the disease etiology in 12 patients with de novo variants in FAR1 all resulting in an amino acid change at position 480 (p.Arg480Cys/His/Leu).
Methods: Following next-generation sequencing and clinical phenotyping, functional characterization was performed in patients' fibroblasts using FAR1 enzyme analysis, FAR1 immunoblotting/immunofluorescence, and lipidomics.
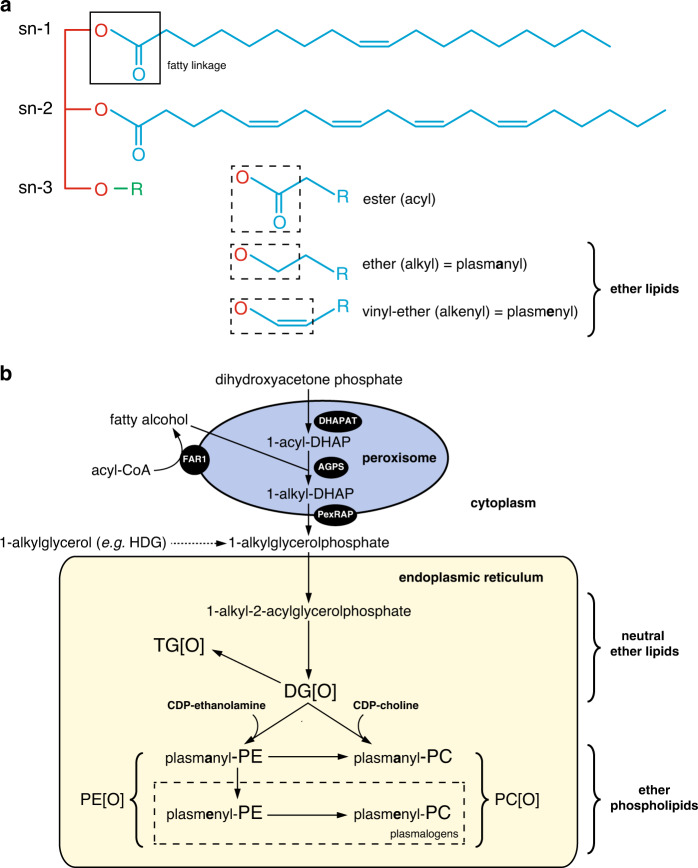
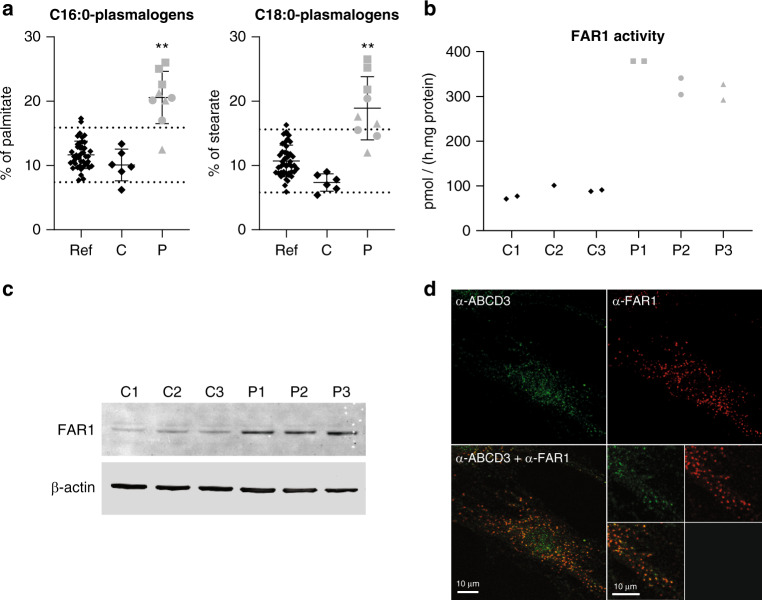
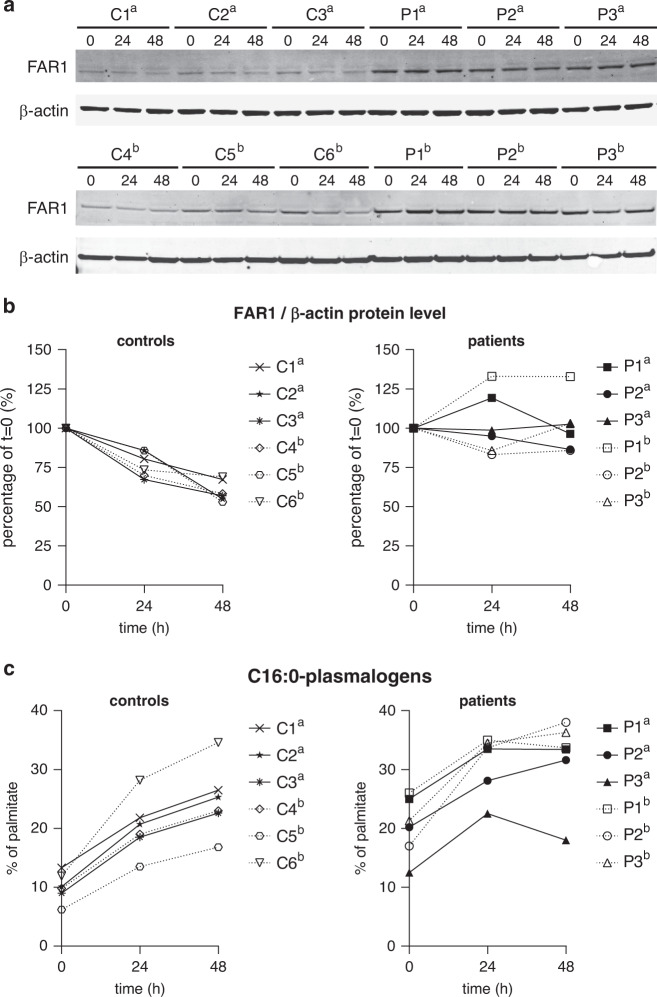
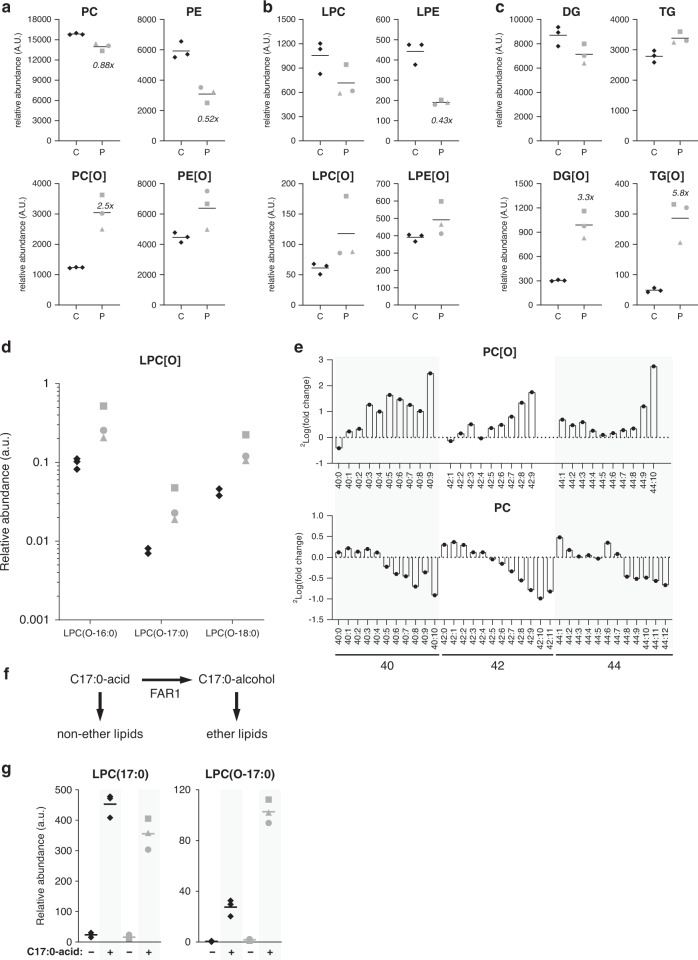
Results: All patients had spastic paraparesis and bilateral congenital/juvenile cataracts, in most combined with speech and gross motor developmental delay and truncal hypotonia. FAR1 deficiency caused by biallelic variants results in defective ether lipid synthesis and plasmalogen deficiency. In contrast, patients' fibroblasts with the de novo FAR1 variants showed elevated plasmalogen levels. Further functional studies in fibroblasts showed that these variants cause a disruption of the plasmalogen-dependent feedback regulation of FAR1 protein levels leading to uncontrolled ether lipid production.
Conclusion: Heterozygous de novo variants affecting the Arg480 residue of FAR1 lead to an autosomal dominant disorder with a different disease mechanism than that of recessive FAR1 deficiency and a diametrically opposed biochemical phenotype. Our findings show that for patients with spastic paraparesis and bilateral cataracts, FAR1 should be considered as a candidate gene and added to gene panels for hereditary spastic paraplegia, cerebral palsy, and juvenile cataracts.
Conflict of interest statement
K.M., A.B., F.M., M.J.G.S. are employees of GeneDx, Inc. A.V.D. is an advisory board member for ProQr. The other authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
