

Multiple wheat genomes reveal global variation in modern breeding
- PMID: 33239791
- PMCID: PMC7759465
- DOI: 10.1038/s41586-020-2961-x
Multiple wheat genomes reveal global variation in modern breeding
Abstract
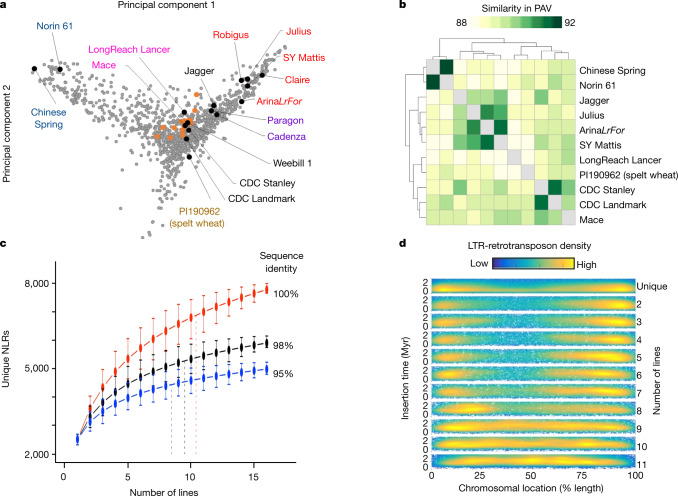
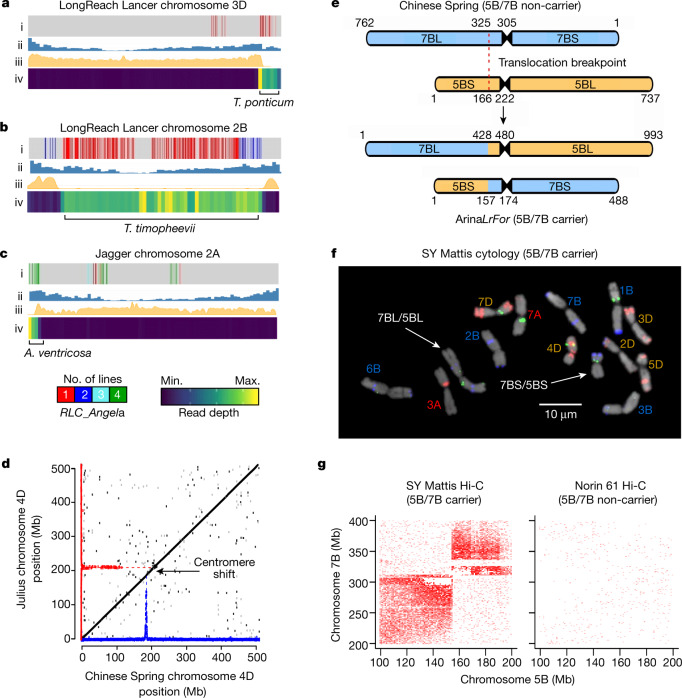
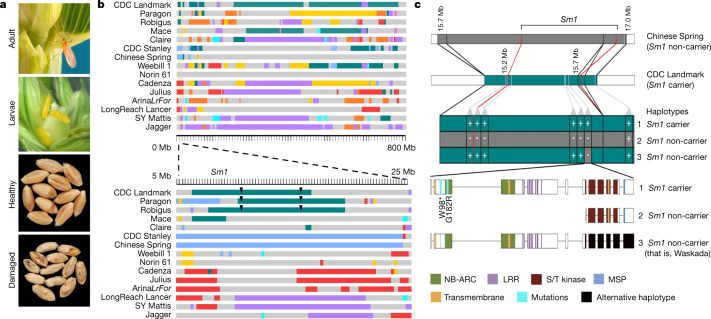
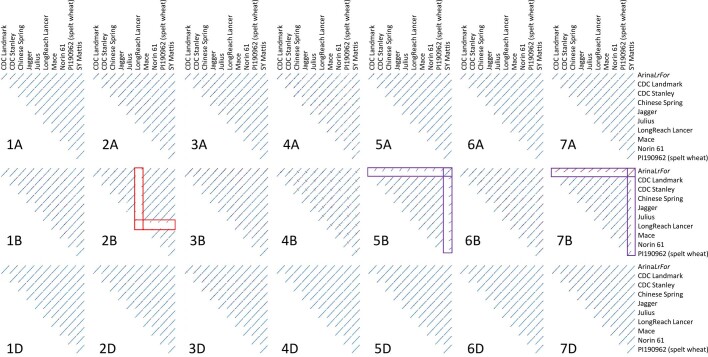
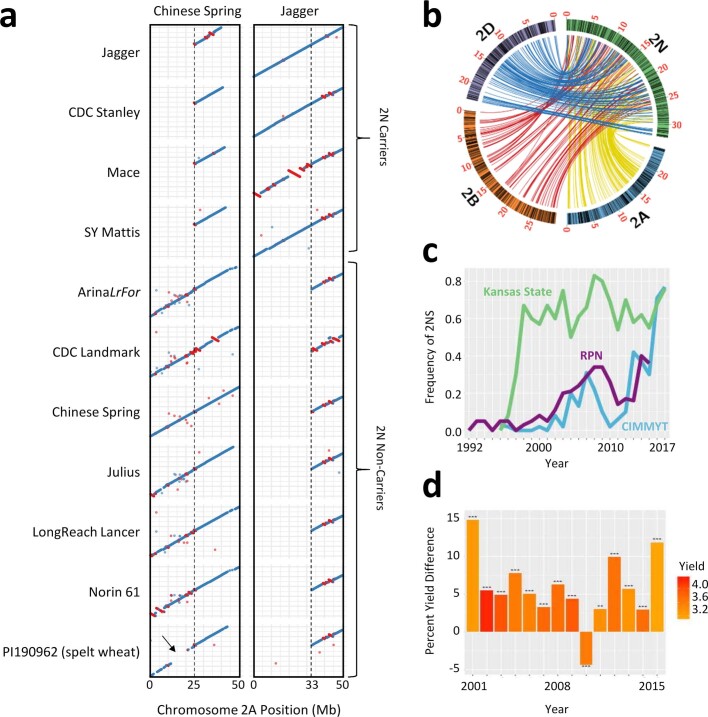
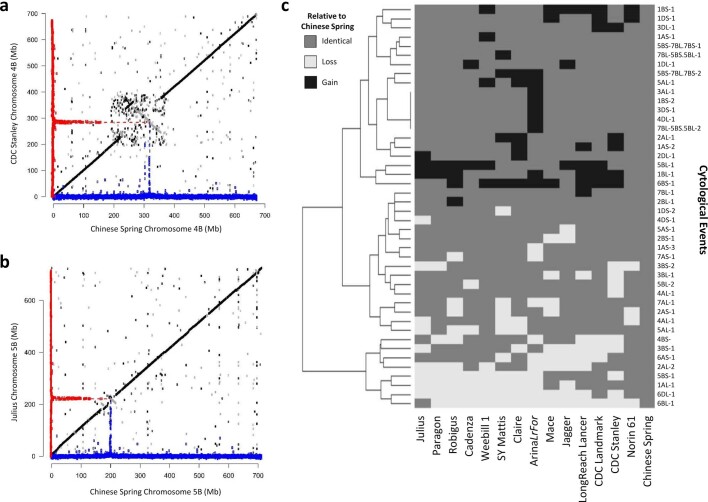
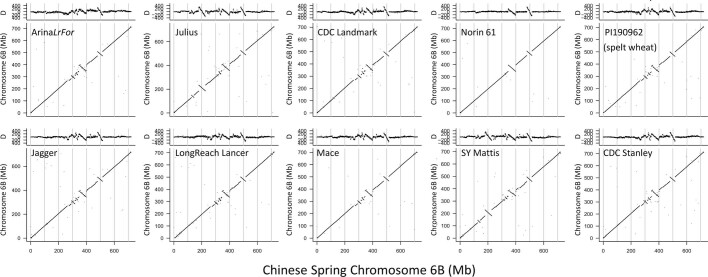
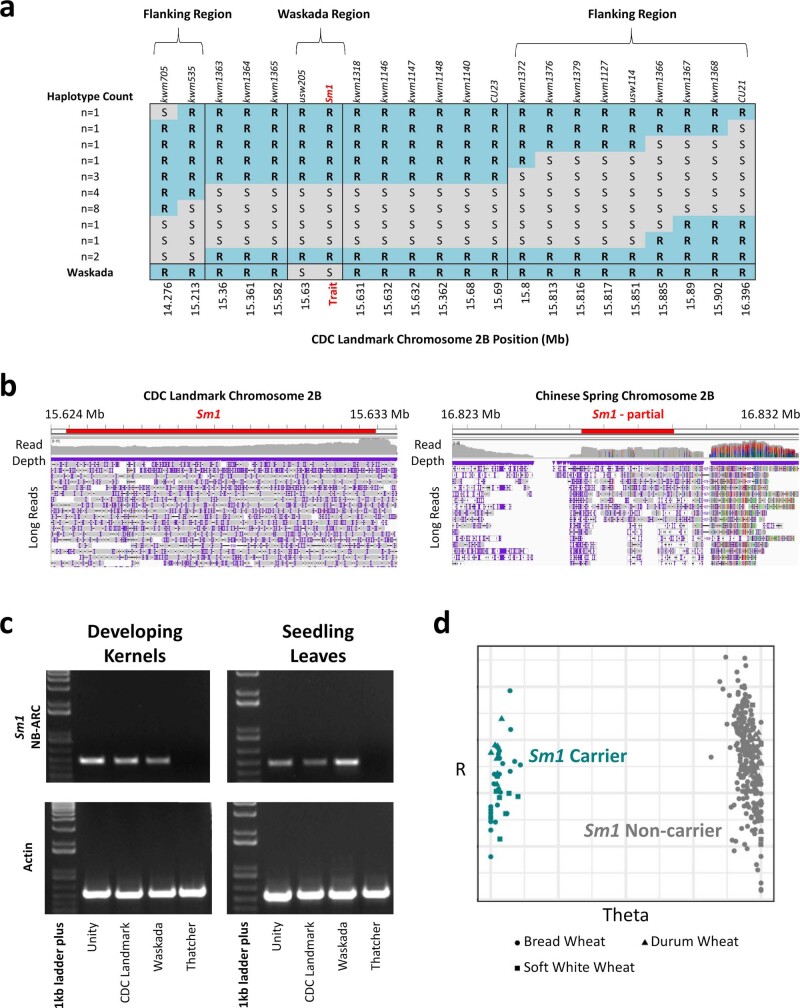
Advances in genomics have expedited the improvement of several agriculturally important crops but similar efforts in wheat (Triticum spp.) have been more challenging. This is largely owing to the size and complexity of the wheat genome1, and the lack of genome-assembly data for multiple wheat lines2,3. Here we generated ten chromosome pseudomolecule and five scaffold assemblies of hexaploid wheat to explore the genomic diversity among wheat lines from global breeding programs. Comparative analysis revealed extensive structural rearrangements, introgressions from wild relatives and differences in gene content resulting from complex breeding histories aimed at improving adaptation to diverse environments, grain yield and quality, and resistance to stresses4,5. We provide examples outlining the utility of these genomes, including a detailed multi-genome-derived nucleotide-binding leucine-rich repeat protein repertoire involved in disease resistance and the characterization of Sm16, a gene associated with insect resistance. These genome assemblies will provide a basis for functional gene discovery and breeding to deliver the next generation of modern wheat cultivars.
Conflict of interest statement
The authors declare no competing interests.
Figures
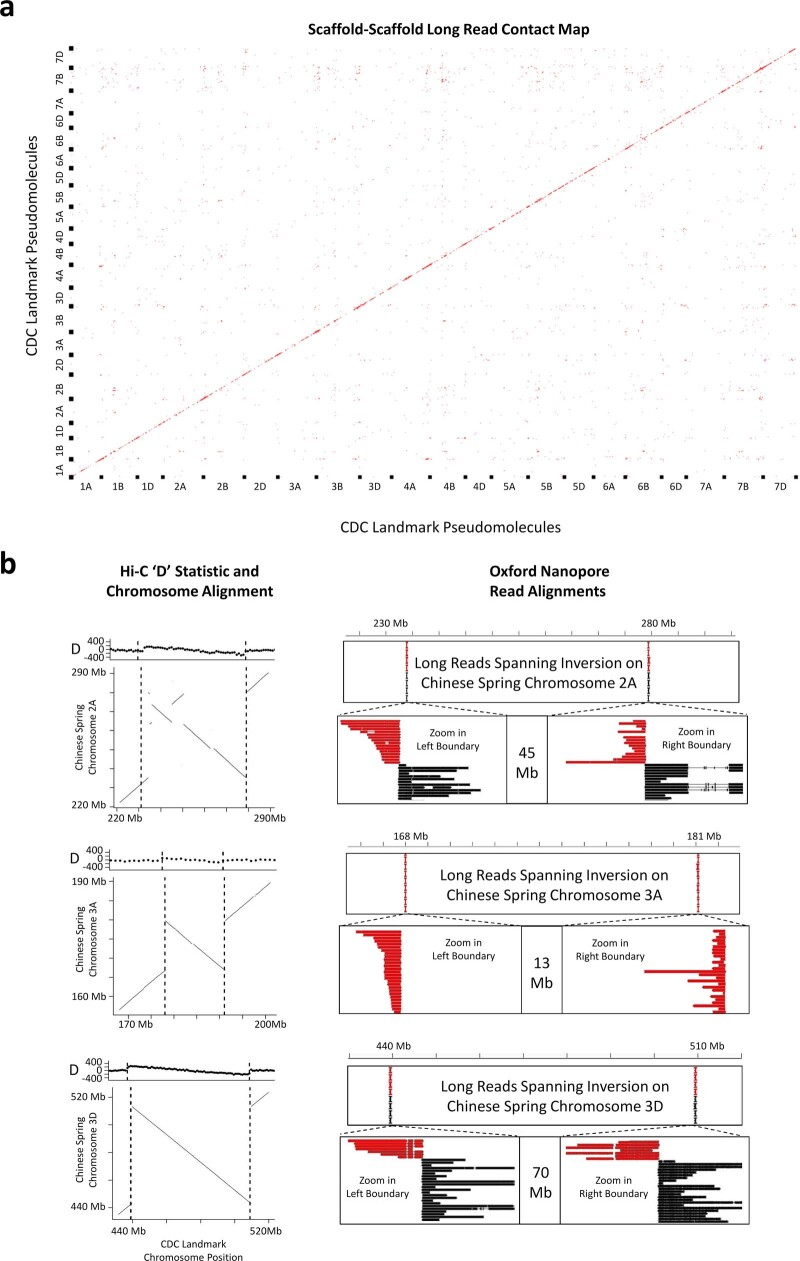
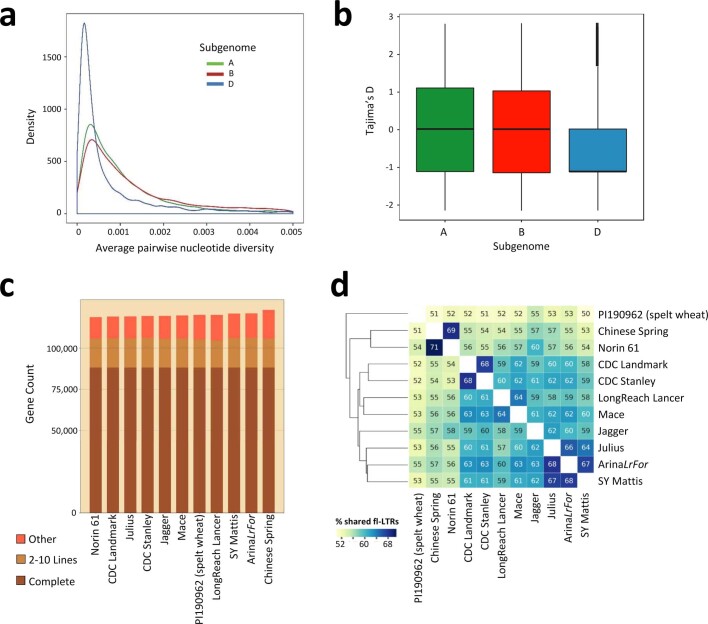
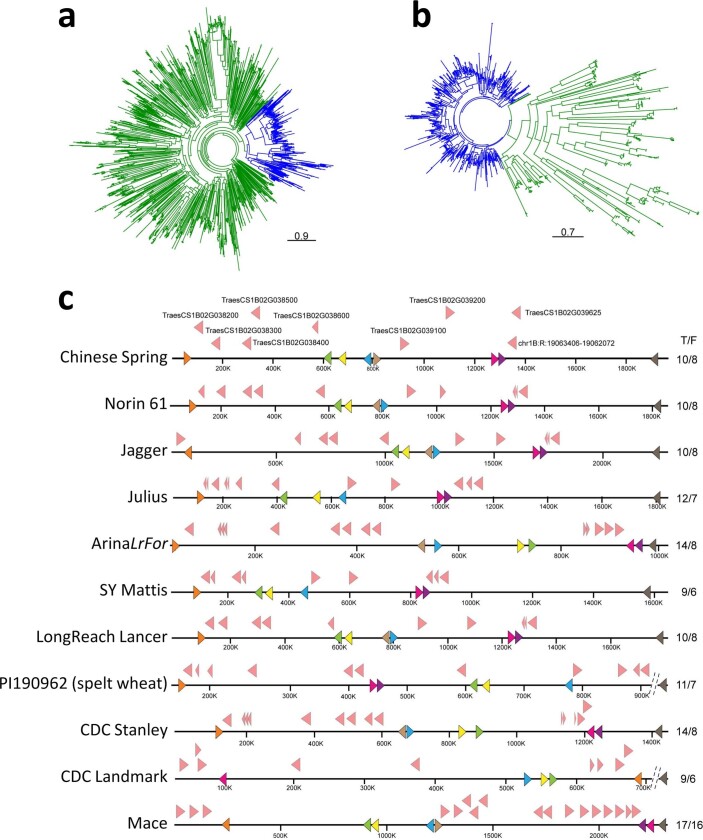
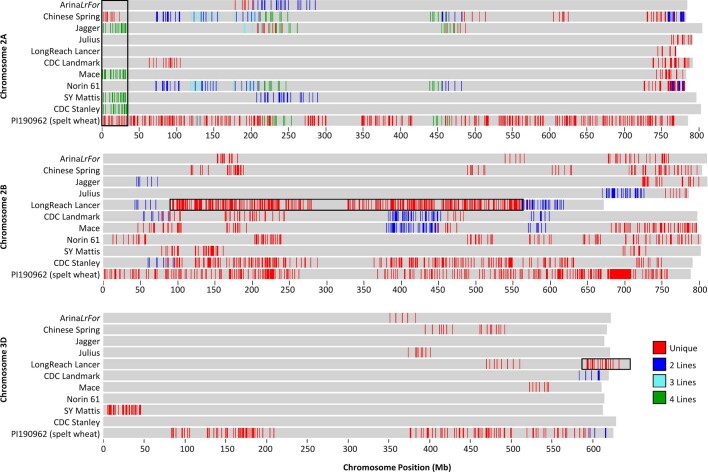

References
-
- The International Wheat Genome Sequencing Consortium Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science. 2018;361:eaar7191. - PubMed
-
- Montenegro JD, et al. The pangenome of hexaploid bread wheat. Plant J. 2017;90:1007–1013. - PubMed
-
- International Wheat Genome Sequencing Consortium (IWGSC) A chromosome-based draft sequence of the hexaploid bread wheat (Triticum aestivum) genome. Science. 2014;345:1251788. - PubMed
-
- He F, et al. Exome sequencing highlights the role of wild-relative introgression in shaping the adaptive landscape of the wheat genome. Nat. Genet. 2019;51:1194–1194. - PubMed
-
- Pont C, et al. Tracing the ancestry of modern bread wheats. Nat. Genet. 2019;51:905–911. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
