

Discoveries in structure and physiology of mechanically activated ion channels
- PMID: 33239794
- PMCID: PMC8477435
- DOI: 10.1038/s41586-020-2933-1
Discoveries in structure and physiology of mechanically activated ion channels
Abstract
The ability to sense physical forces is conserved across all organisms. Cells convert mechanical stimuli into electrical or chemical signals via mechanically activated ion channels. In recent years, the identification of new families of mechanosensitive ion channels-such as PIEZO and OSCA/TMEM63 channels-along with surprising insights into well-studied mechanosensitive channels have driven further developments in the mechanotransduction field. Several well-characterized mechanosensory roles such as touch, blood-pressure sensing and hearing are now linked with primary mechanotransducers. Unanticipated roles of mechanical force sensing continue to be uncovered. Furthermore, high-resolution structures representative of nearly every family of mechanically activated channel described so far have underscored their diversity while advancing our understanding of the biophysical mechanisms of pressure sensing. Here we summarize recent discoveries in the physiology and structures of known mechanically activated ion channel families and discuss their implications for understanding the mechanisms of mechanical force sensing.
Conflict of interest statement
Figures
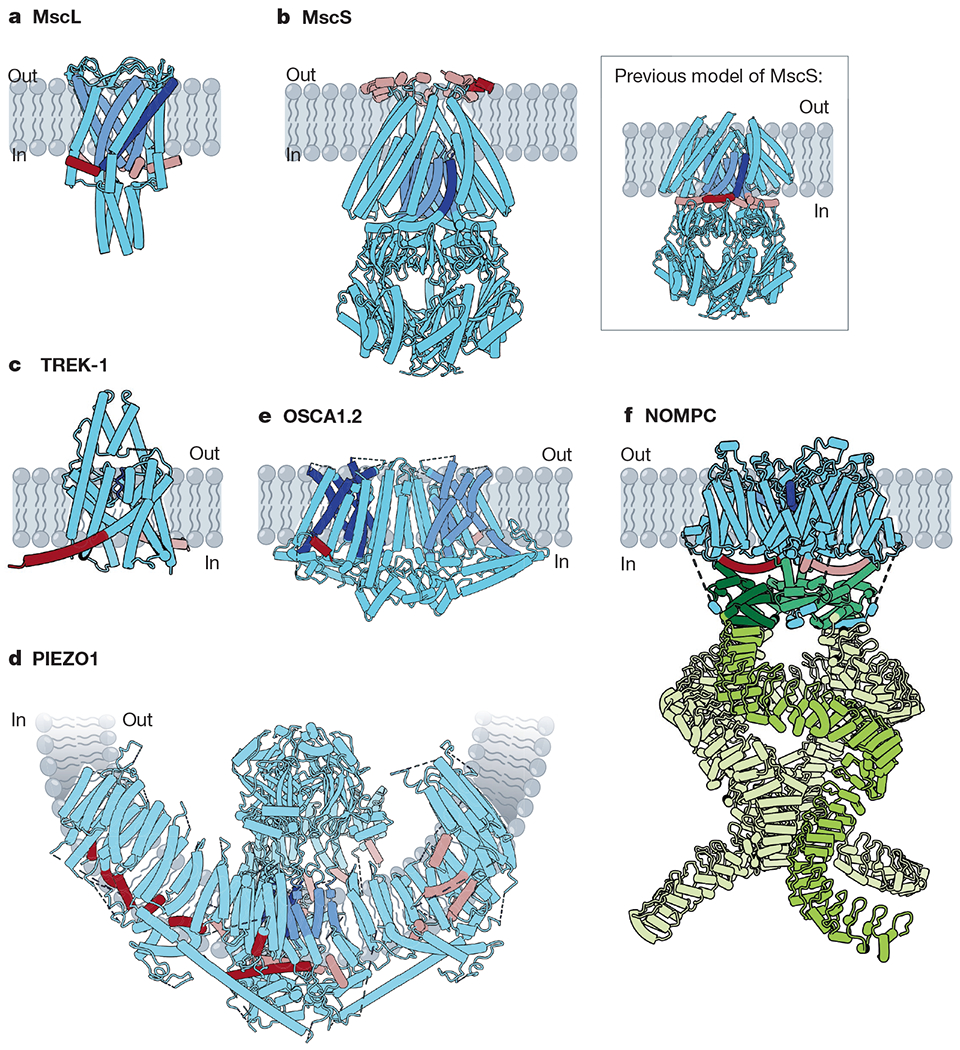
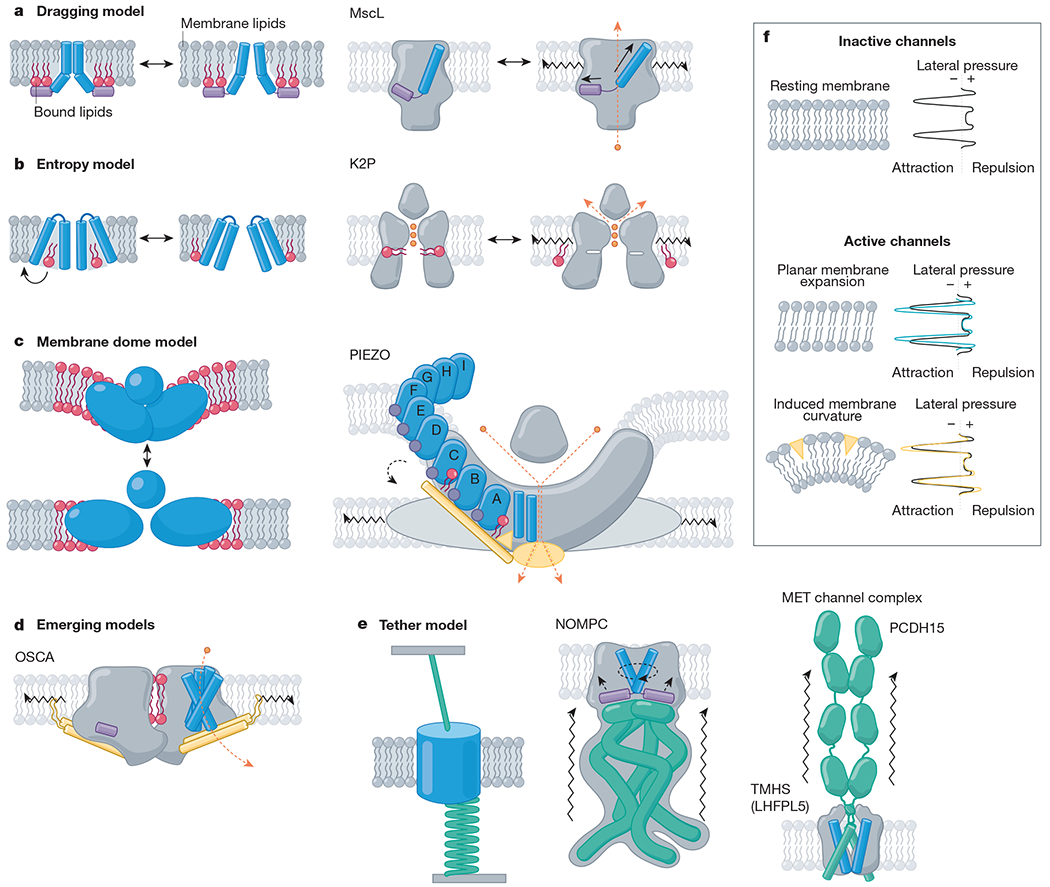
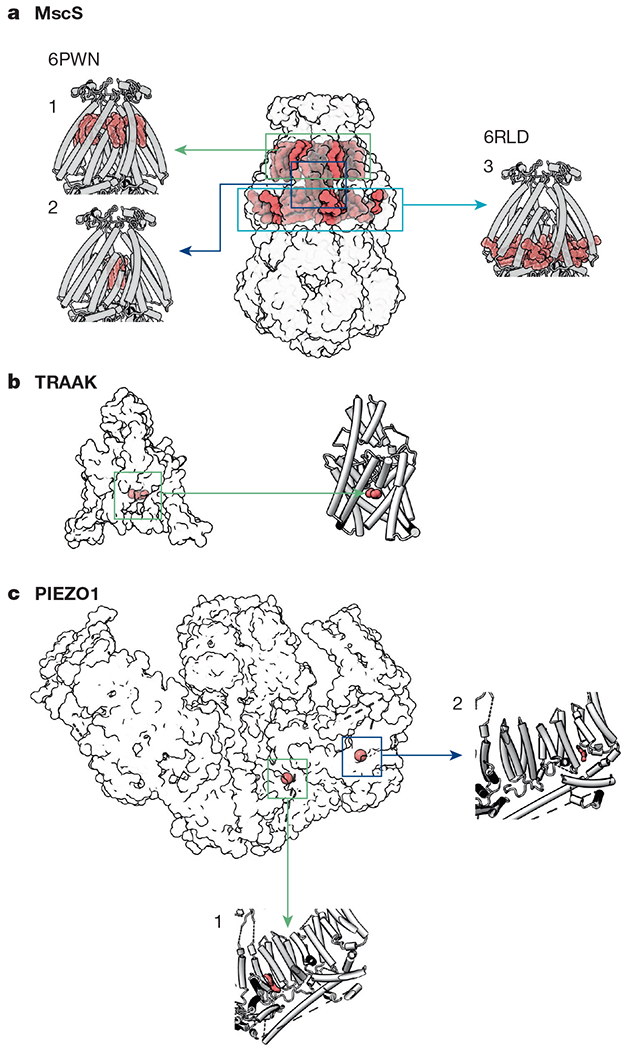
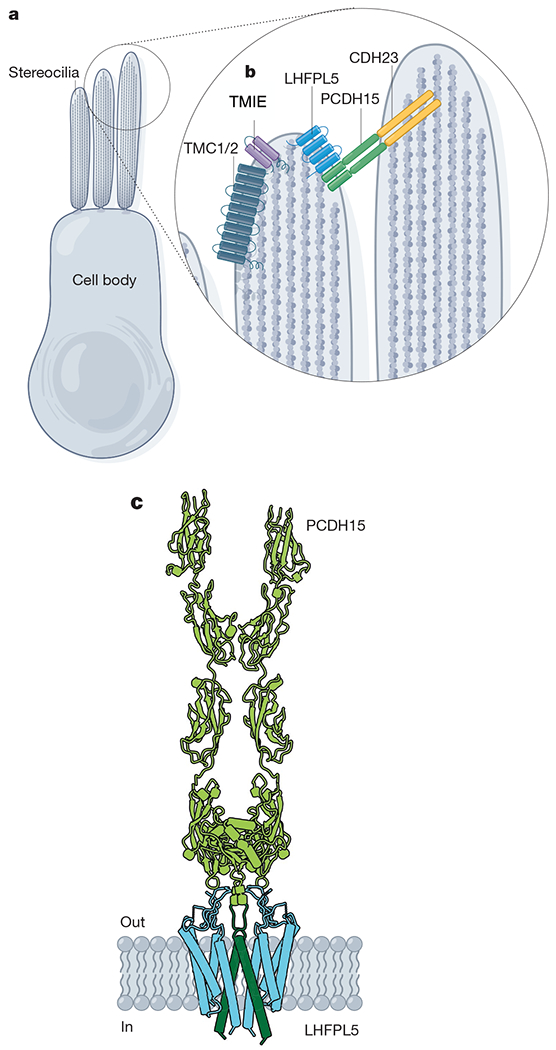
References
-
- Arnadóttir J & Chalfie M Eukaryotic mechanosensitive channels. Annu. Rev. Biophys 39, 111–137 (2010). - PubMed
-
- Cox CD, Bavi N & Martinac B Bacterial mechanosensors. Annu. Rev. Physiol 80, 71–93 (2018). - PubMed
-
- Chalfie M Neurosensory mechanotransduction. Nat. Rev. Mol. Cell Biol 10, 44–52 (2009). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
