

De Novo Genome Assembly of the Japanese Wheat Cultivar Norin 61 Highlights Functional Variation in Flowering Time and Fusarium-Resistant Genes in East Asian Genotypes
- PMID: 33244607
- PMCID: PMC7991897
- DOI: 10.1093/pcp/pcaa152
De Novo Genome Assembly of the Japanese Wheat Cultivar Norin 61 Highlights Functional Variation in Flowering Time and Fusarium-Resistant Genes in East Asian Genotypes
Abstract
Bread wheat is a major crop that has long been the focus of basic and breeding research. Assembly of its genome has been difficult because of its large size and allohexaploid nature (AABBDD genome). Following the first reported assembly of the genome of the experimental strain Chinese Spring (CS), the 10+ Wheat Genomes Project was launched to produce multiple assemblies of worldwide modern cultivars. The only Asian cultivar in the project is Norin 61, a representative Japanese cultivar adapted to grow across a broad latitudinal range, mostly characterized by a wet climate and a short growing season. Here, we characterize the key aspects of its chromosome-scale genome assembly spanning 15 Gb with a raw scaffold N50 of 22 Mb. Analysis of the repetitive elements identified chromosomal regions unique to Norin 61 that encompass a tandem array of the pathogenesis-related 13 family. We report novel copy-number variations in the B homeolog of the florigen gene FT1/VRN3, pseudogenization of its D homeolog and the association of its A homeologous alleles with the spring/winter growth habit. Furthermore, the Norin 61 genome carries typical East Asian functional variants different from CS, ranging from a single nucleotide to multi-Mb scale. Examples of such variation are the Fhb1 locus, which confers Fusarium head-blight resistance, Ppd-D1a, which confers early flowering, Glu-D1f for Asian noodle quality and Rht-D1b, which introduced semi-dwarfism during the green revolution. The adoption of Norin 61 as a reference assembly for functional and evolutionary studies will enable comprehensive characterization of the underexploited Asian bread wheat diversity.
Keywords: Adaptation; Asian germplasm; Bread wheat; Genome assembly; Norin 61; Polyploidy.
� The Author(s) 2021. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists.
Figures

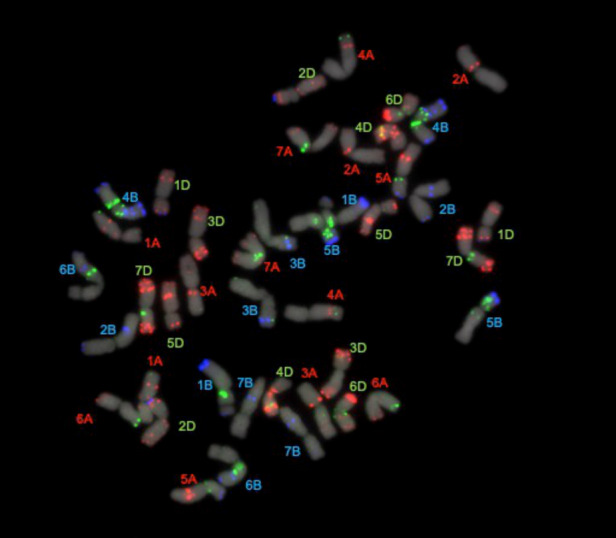
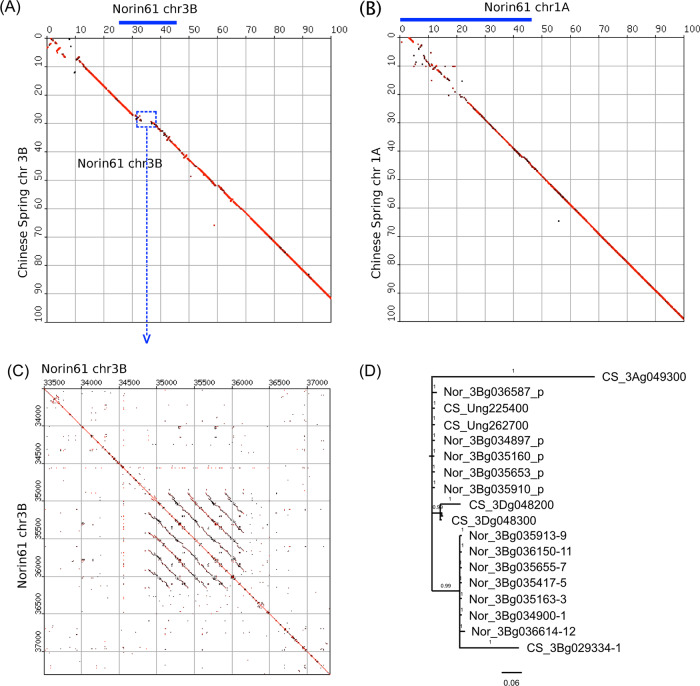
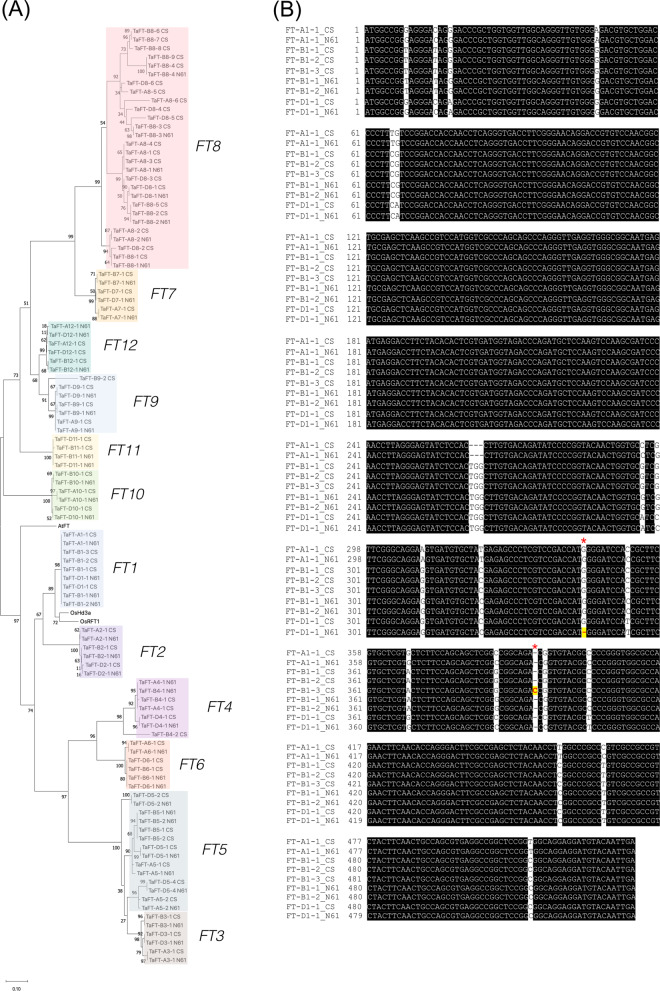



References
-
- Ammerman A.J., Cavalli-Sforza L.L. (1971) Measuring the rate of spread of early farming in Europe. Man 6: 674.
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
