

Single-cell RNA sequencing and lipidomics reveal cell and lipid dynamics of fat infiltration in skeletal muscle
- PMID: 33244879
- PMCID: PMC7890272
- DOI: 10.1002/jcsm.12643
Single-cell RNA sequencing and lipidomics reveal cell and lipid dynamics of fat infiltration in skeletal muscle
Abstract
Background: Ageing is accompanied by sarcopenia and intramuscular fat (IMAT) infiltration. In skeletal muscle, fat infiltration is a common feature in several myopathies and is associated with muscular dysfunction and insulin resistance. However, the cellular origin and lipidomic and transcriptomic changes during fat infiltration in skeletal muscle remain unclear.
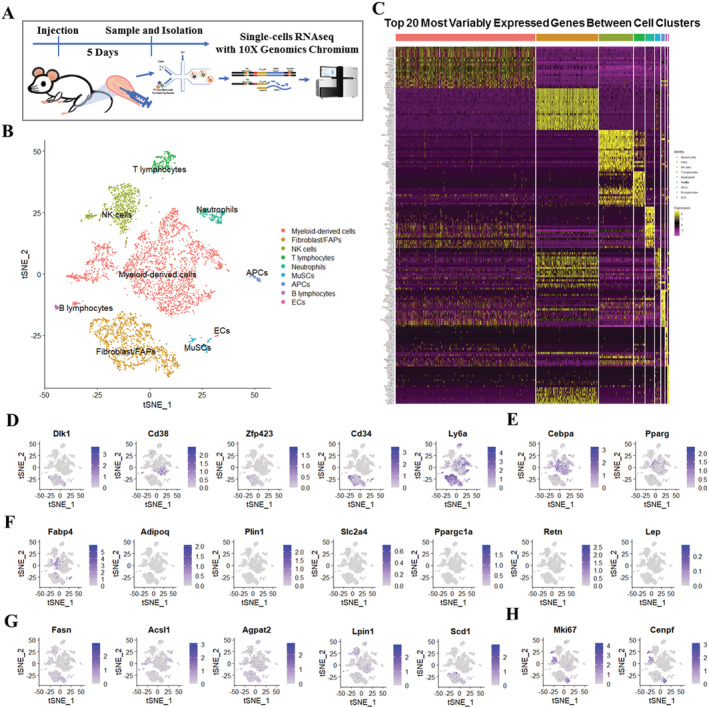
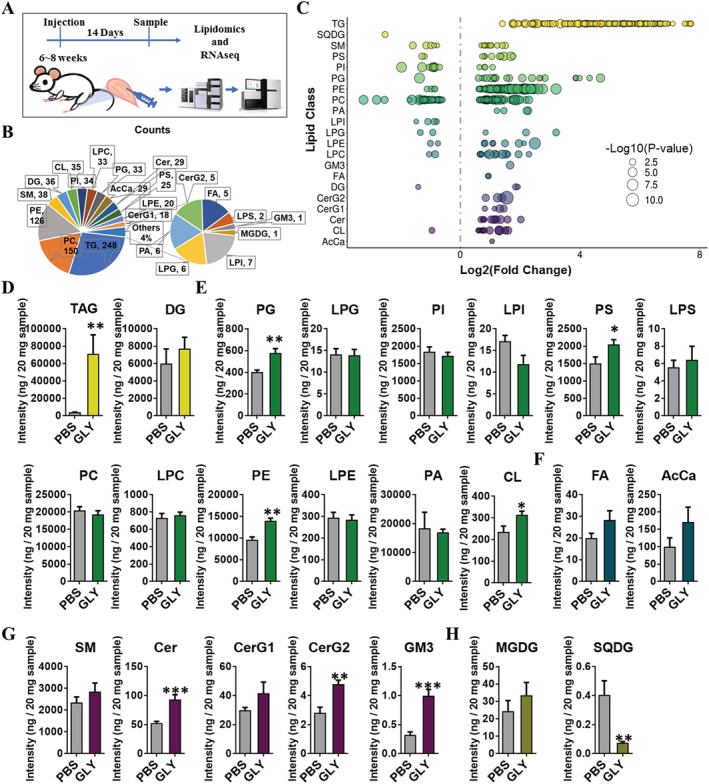
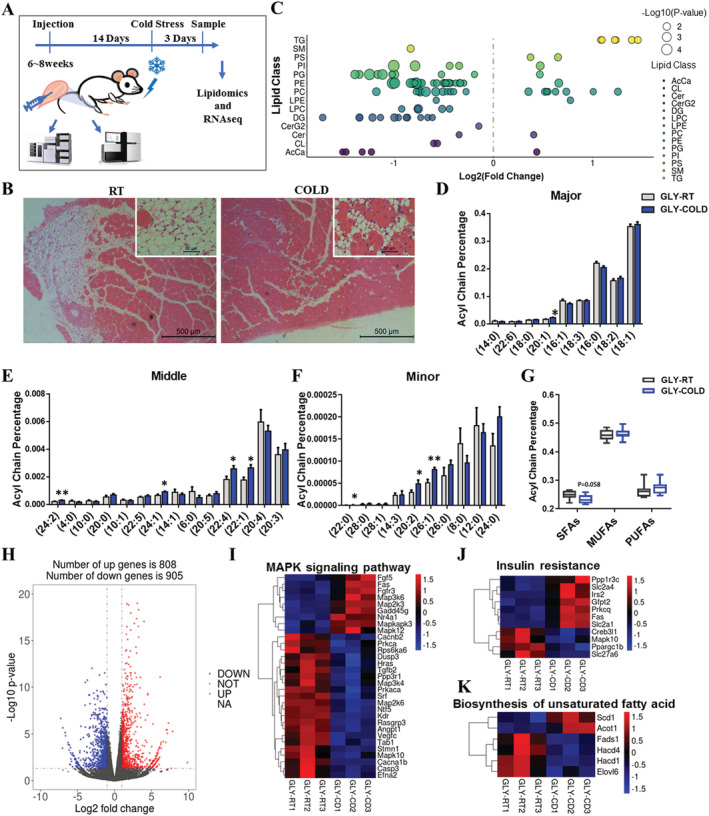
Methods: In the current study, we generated a high IMAT-infiltrated skeletal muscle model by glycerol (GLY) injection. Single-cell RNA sequencing and lineage tracing were performed on GLY-injured skeletal muscle at 5 days post-injection (DPI) to identify the cell origins and dynamics. Lipidomics and RNA sequencing were performed on IMAT-infiltrated skeletal muscle at 14 DPI (or 17 DPI for the cold treatment) to analyse alterations of lipid compositions and gene expression levels.
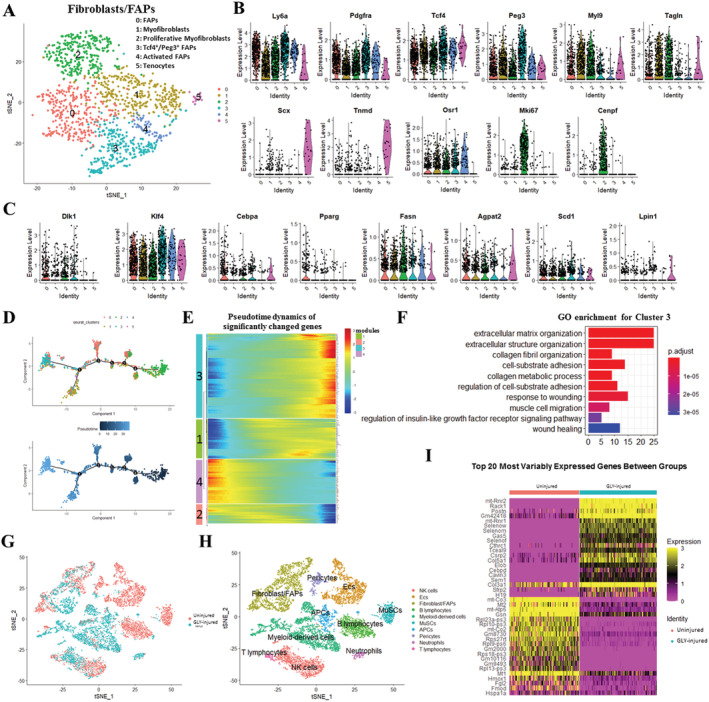
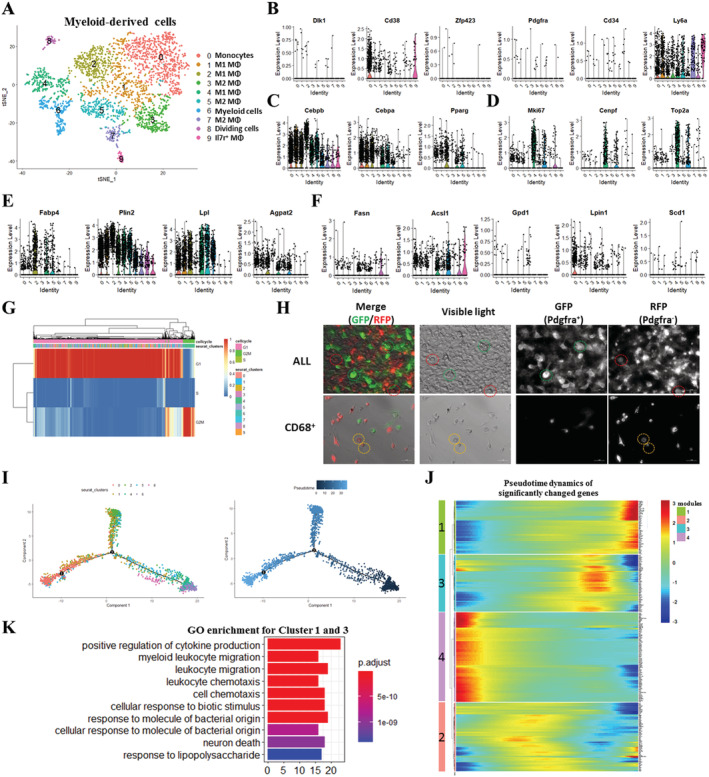
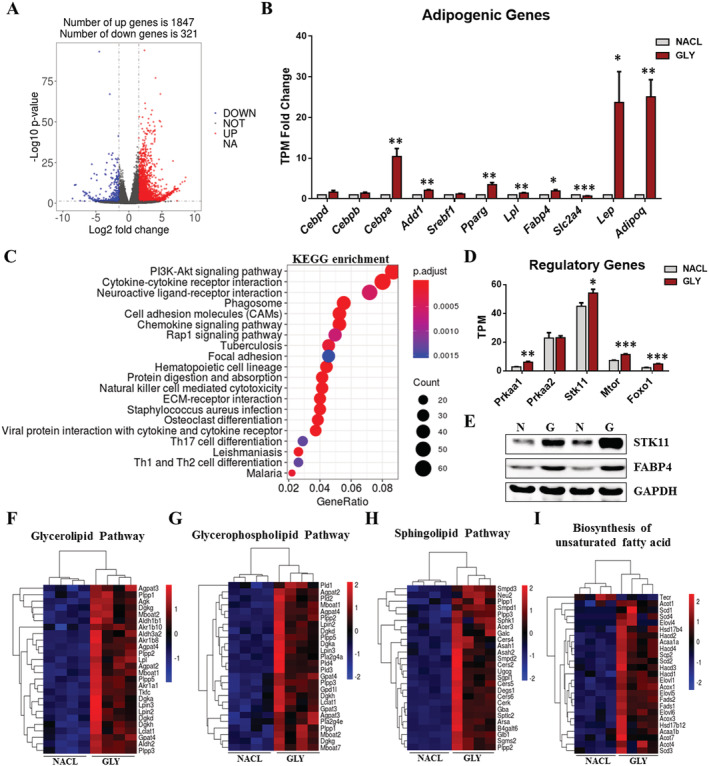
Results: We identified nine distinct major clusters including myeloid-derived cells (52.13%), fibroblast/fibro/adipogenic progenitors (FAPs) (23.24%), and skeletal muscle stem cells (2.02%) in GLY-injured skeletal muscle. Clustering and pseudotemporal trajectories revealed six subpopulations in fibroblast/FAPs and 10 subclusters in myeloid-derived cells. A subpopulation of myeloid-derived cells expressing adipocyte-enriched genes and Pdgfra- /Cd68+ cells displayed lipid droplets upon adipogenic induction, indicating their adipogenic potential. Lipidomic analysis revealed the changes of overall lipid classes composition (e.g. triglycerides (TAGs) increased by 19.3 times, P = 0.0098; sulfoquinovosyl diacylglycerol decreased by 83%, P = 0.0056) and in the distribution of lipids [e.g. TAGs (18:2/18:2/22:6) increased by 181.6 times, P = 0.021] between GLY-group and saline control. RNA-seq revealed 1847 up-regulated genes and 321 down-regulated genes and significant changes in lipid metabolism-related pathways (e.g. glycerolipid pathway and glycerophospholipid pathway) in our model of GLY-injured skeletal muscle. Notably, short-term cold exposure altered fatty acid composition (e.g. saturated fatty acid decreased by 6.4%, P = 0.058) in fat-infiltrated muscles through directly affecting lipid metabolism pathways including PI3K-AKT and MAPK signalling pathway.
Conclusions: Our results showed that a subpopulation of myeloid-derived cells may contribute to IMAT infiltration. GLY-induced IMAT infiltration changed the lipid composition and gene expression profiles. Short-term cold exposure might regulate lipid metabolism and its related signalling pathways in fat-infiltrated muscle. Our study provides a comprehensive resource describing the molecular signature of fat infiltration in skeletal muscle.
Keywords: Fat infiltration; Intramuscular fat; Lipidomics; Muscle wasting; Sarcopenia; Single-cell RNA sequencing; Skeletal muscle.
© 2020 The Authors. Journal of Cachexia, Sarcopenia and Muscle published by John Wiley & Sons Ltd on behalf of Society on Sarcopenia, Cachexia and Wasting Disorders.
Conflict of interest statement
None declared.
Figures
References
-
- Brioche T, Pagano AF, Py G, Chopard A. Muscle wasting and aging: experimental models, fatty infiltrations, and prevention. Mol Aspects Med 2016;50:56–87. - PubMed
-
- Greco AV, Mingrone G, Giancaterini A, Manco M, Morroni M, Cinti S, et al. Insulin resistance in morbid obesity: reversal with intramyocellular fat depletion. Diabetes 2002;51:144–151. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
