

4D nucleome modeling
- PMID: 33253996
- PMCID: PMC8098745
- DOI: 10.1016/j.gde.2020.10.004
4D nucleome modeling
Abstract
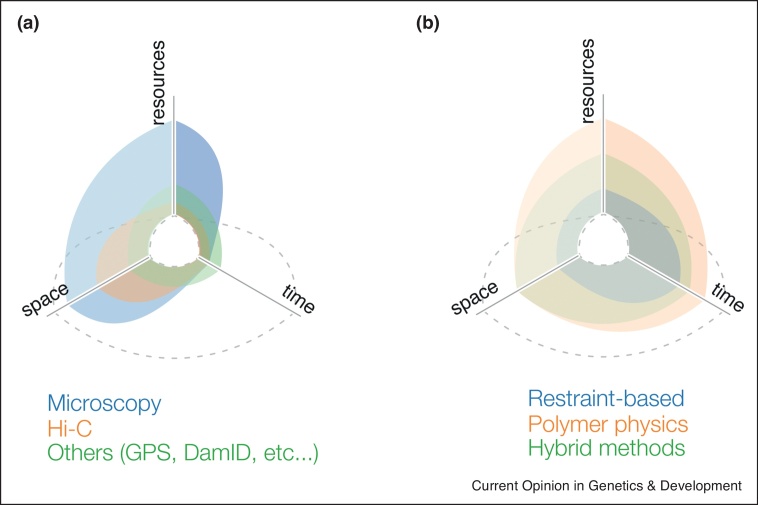
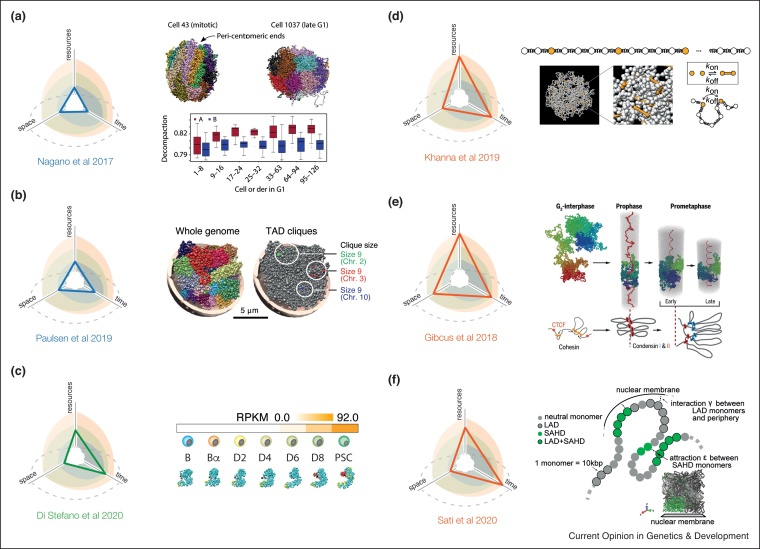
The intrinsic dynamic nature of chromosomes is emerging as a fundamental component in regulating DNA transcription, replication, and damage-repair among other nuclear functions. With this increased awareness, reinforced over the last ten years, many new experimental techniques, mainly based on microscopy and chromosome conformation capture, have been introduced to study the genome in space and time. Owing to the increasing complexity of these cutting-edge techniques, computational approaches have become of paramount importance to interpret, contextualize, and complement such experiments with new insights. Hence, it is becoming crucial for experimental biologists to have a clear understanding of the diverse theoretical modeling approaches available and the biological information each of them can provide.
Copyright © 2020 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
