

Comparative Genomics and Functional Studies of Wheat BED-NLR Loci
- PMID: 33256067
- PMCID: PMC7761493
- DOI: 10.3390/genes11121406
Comparative Genomics and Functional Studies of Wheat BED-NLR Loci
Abstract
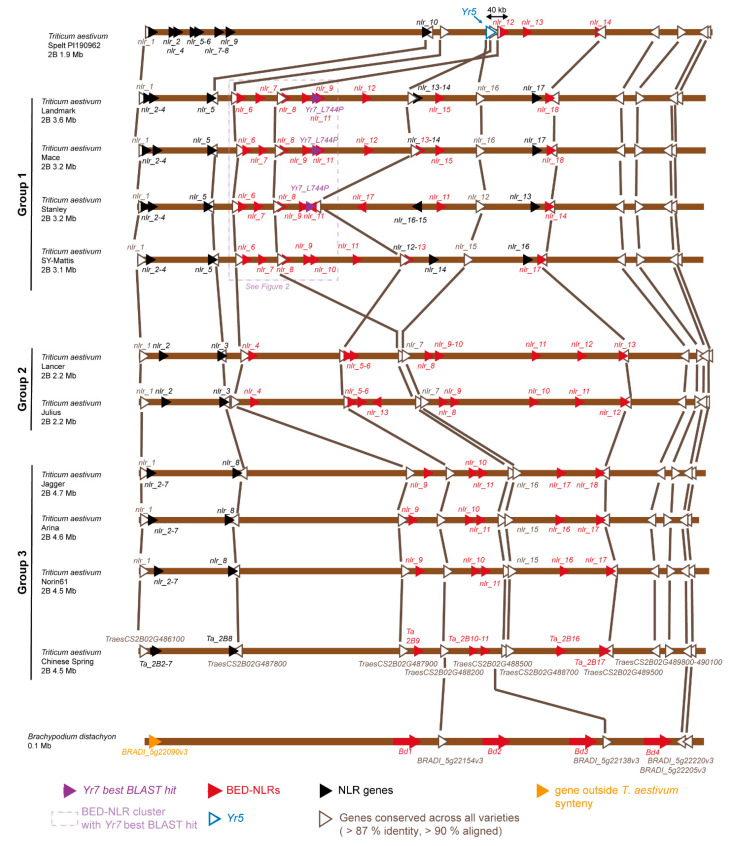
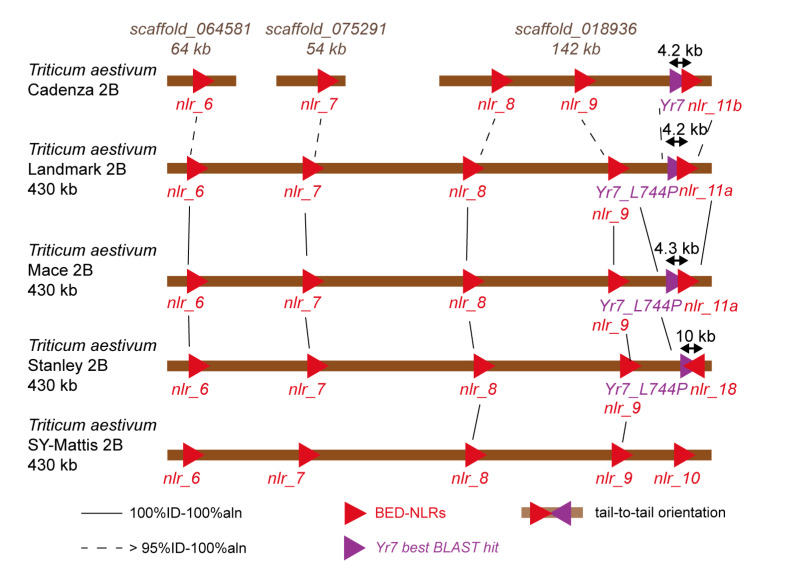
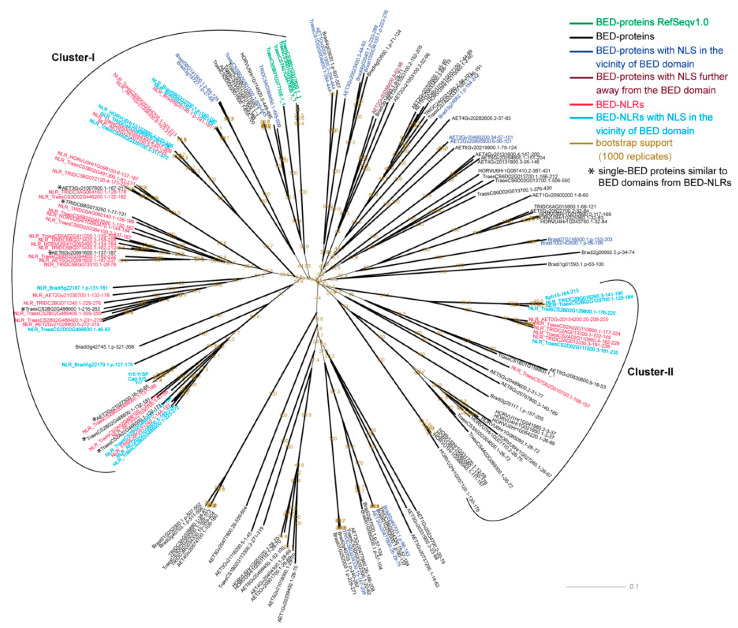
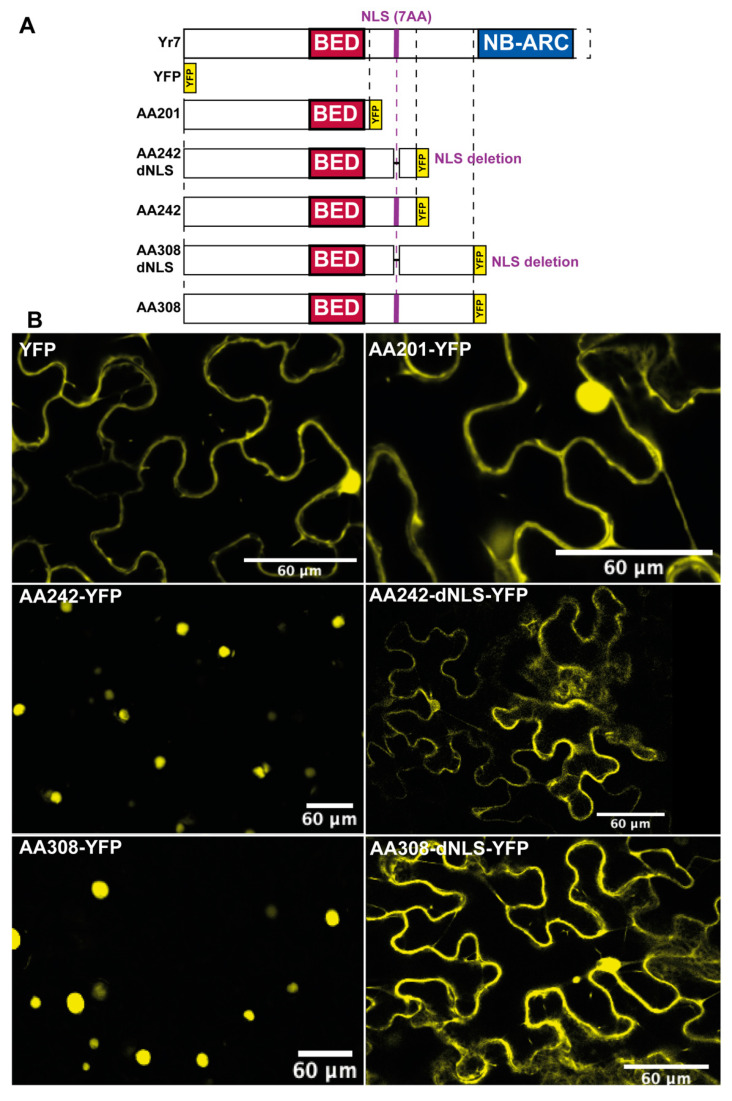
Nucleotide-binding leucine-rich-repeat (LRR) receptors (NLRs) with non-canonical integrated domains (NLR-IDs) are widespread in plant genomes. Zinc-finger BED (named after the Drosophila proteins Boundary Element-Associated Factor and DNA Replication-related Element binding Factor, named BED hereafter) are among the most frequently found IDs. Five BED-NLRs conferring resistance against bacterial and fungal pathogens have been characterized. However, it is unknown whether BED-NLRs function in a manner similar to other NLR-IDs. Here, we used chromosome-level assemblies of wheat to explore the Yr7 and Yr5a genomic regions and show that, unlike known NLR-ID loci, there is no evidence for a NLR-partner in their vicinity. Using neighbor-network analyses, we observed that BED domains from BED-NLRs share more similarities with BED domains from single-BED proteins and from BED-containing proteins harboring domains that are conserved in transposases. We identified a nuclear localization signal (NLS) in Yr7, Yr5, and the other characterized BED-NLRs. We thus propose that this is a feature of BED-NLRs that confer resistance to plant pathogens. We show that the NLS was functional in truncated versions of the Yr7 protein when expressed in N. benthamiana. We did not observe cell-death upon the overexpression of Yr7 full-length, truncated, and 'MHD' variants in N. benthamiana. This suggests that either this system is not suitable to study BED-NLR signaling or that BED-NLRs require additional components to trigger cell death. These results define novel future directions to further understand the role of BED domains in BED-NLR mediated resistance.
Keywords: BED-NLR; DUF4413/659(-hAT); NLR; NLR-ID; Yr5; Yr7; integrated domain; plant disease resistance; wheat; zf-BED.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyzes, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
References
-
- Shao Z.Q., Xue J.Y., Wu P., Zhang Y.M., Wu Y., Hang Y.Y., Wang B., Chen J.Q. Large-scale analyses of angiosperm nucleotide-binding site-leucine-rich repeat genes reveal three anciently diverged classes with distinct evolutionary patterns. Plant Physiol. 2016;170:2095–2109. doi: 10.1104/pp.15.01487. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
