

Stress Signaling in Cyanobacteria: A Mechanistic Overview
- PMID: 33256109
- PMCID: PMC7760821
- DOI: 10.3390/life10120312
Stress Signaling in Cyanobacteria: A Mechanistic Overview
Abstract
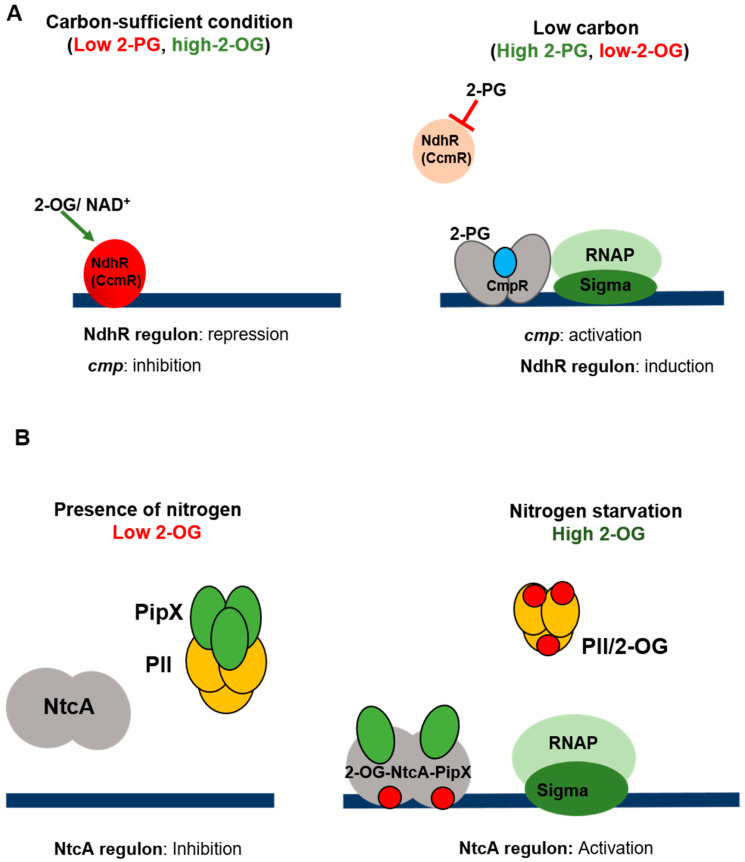
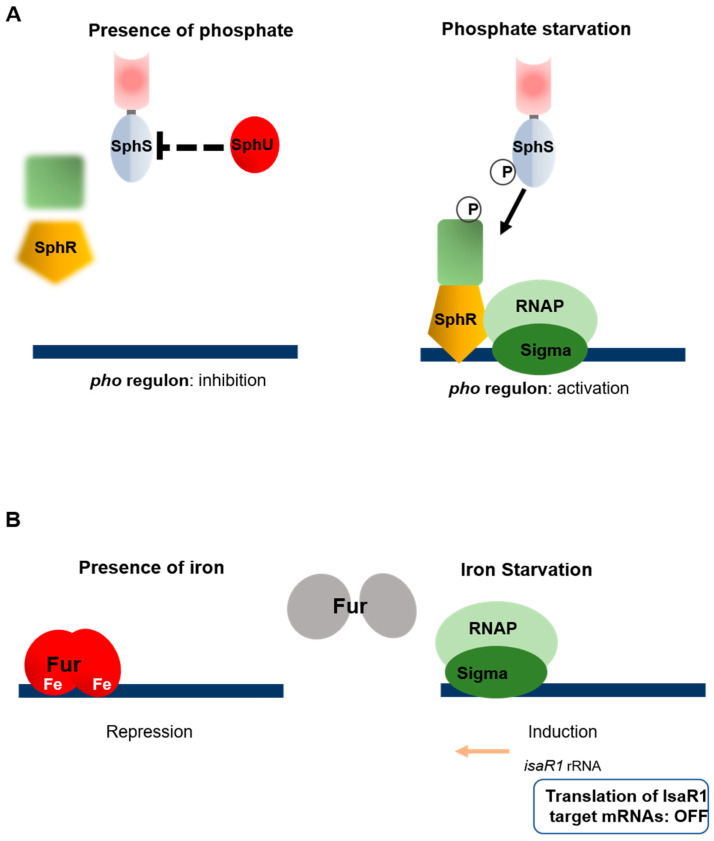
Cyanobacteria are highly diverse, widely distributed photosynthetic bacteria inhabiting various environments ranging from deserts to the cryosphere. Throughout this range of niches, they have to cope with various stresses and kinds of deprivation which threaten their growth and viability. In order to adapt to these stresses and survive, they have developed several global adaptive responses which modulate the patterns of gene expression and the cellular functions at work. Sigma factors, two-component systems, transcriptional regulators and small regulatory RNAs acting either separately or collectively, for example, induce appropriate cyanobacterial stress responses. The aim of this review is to summarize our current knowledge about the diversity of the sensors and regulators involved in the perception and transduction of light, oxidative and thermal stresses, and nutrient starvation responses. The studies discussed here point to the fact that various stresses affecting the photosynthetic capacity are transduced by common mechanisms.
Keywords: cyanobacteria; gene expression; signaling; stress.
Conflict of interest statement
The authors declare that they have no conflicts of interest to declare. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
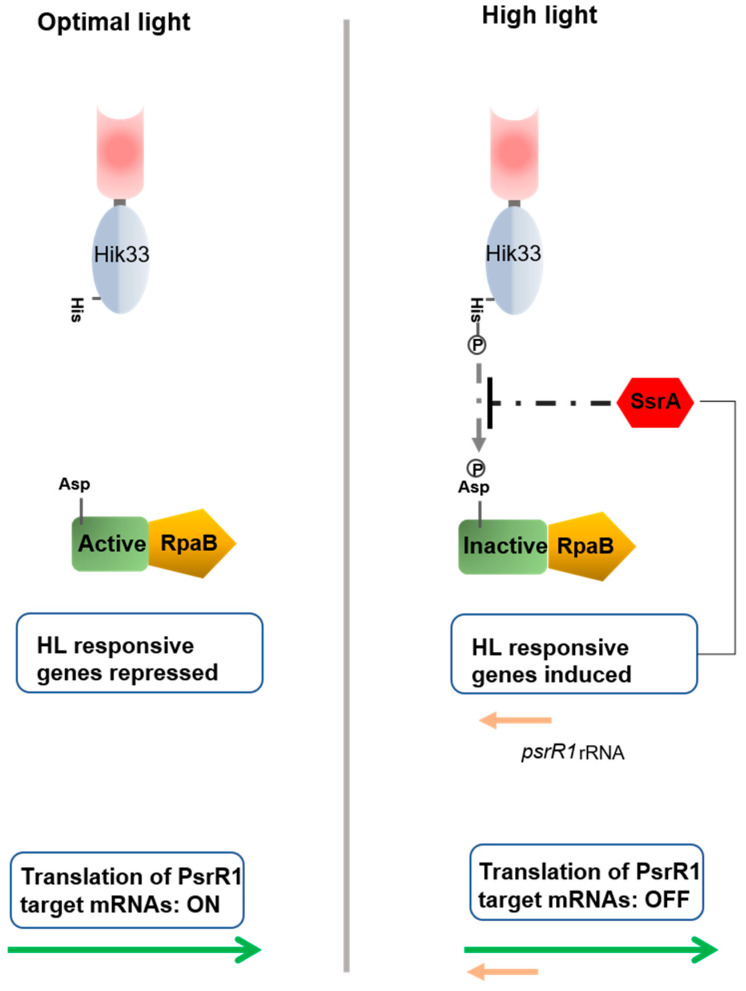
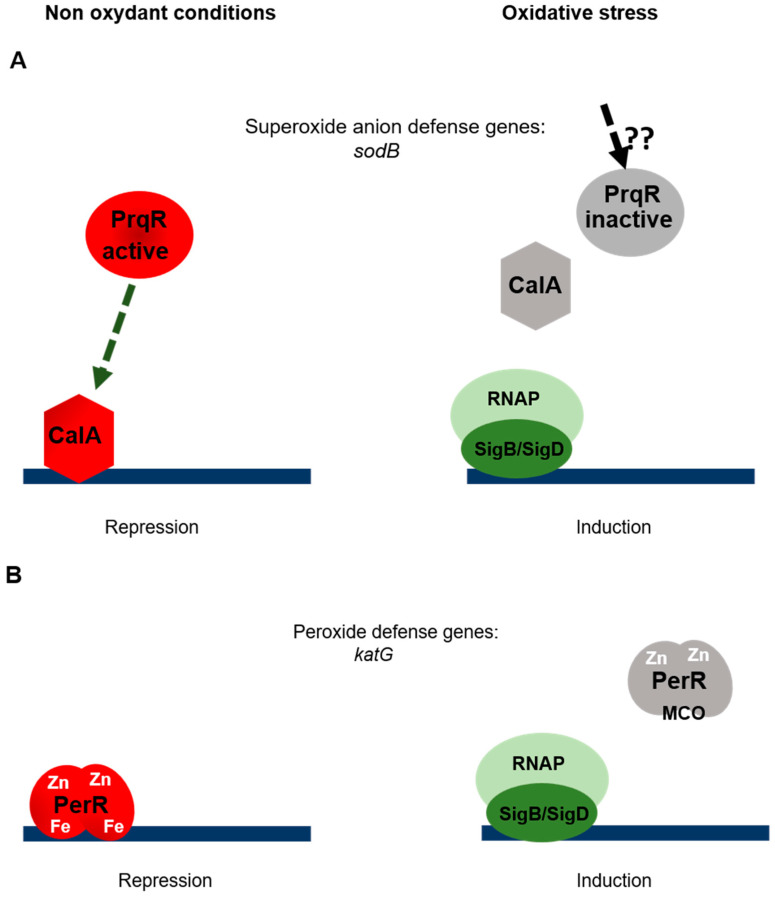
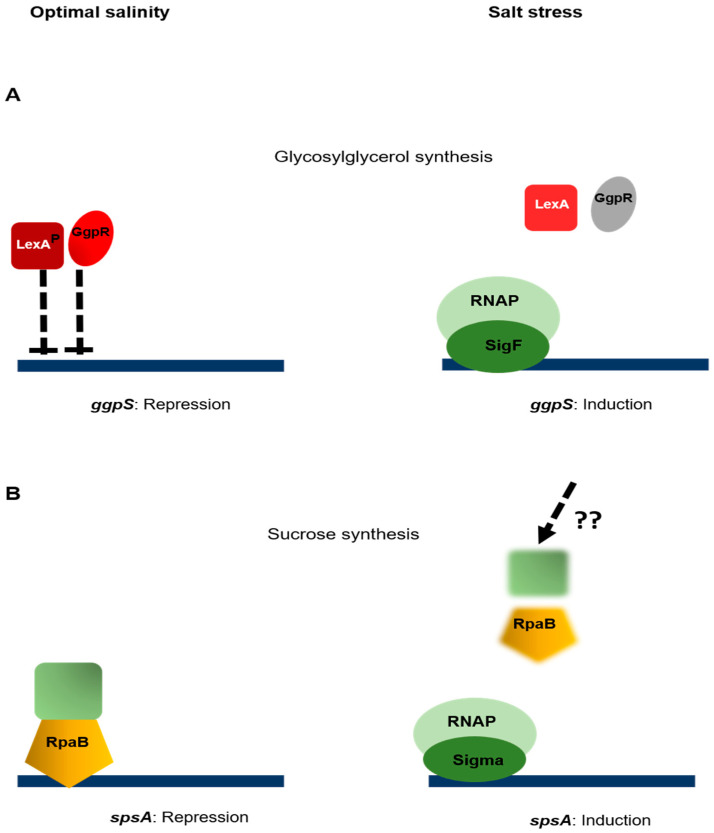
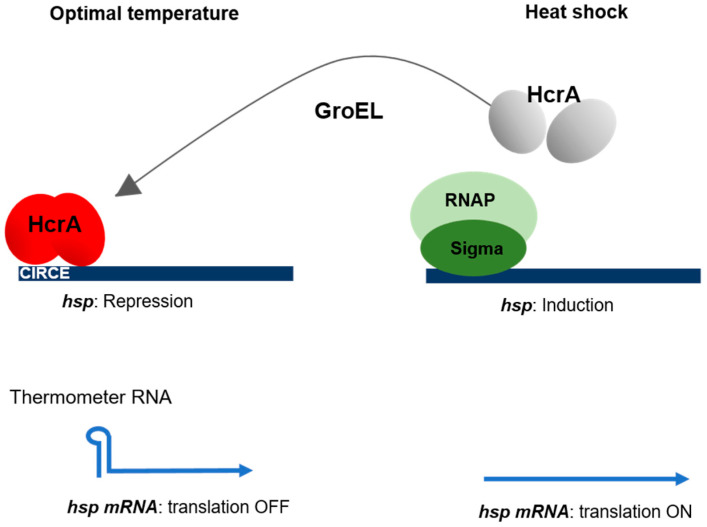
References
-
- Flombaum P., Gallegos J.L., Gordillo R.A., Rincon J., Zabala L.L., Jiao N., Karl D.M., Li W.K., Lomas M.W., Veneziano D., et al. Present and future global distributions of the marine Cyanobacteria Prochlorococcus and Synechococcus. Proc. Natl. Acad. Sci. USA. 2013;110:9824–9829. doi: 10.1073/pnas.1307701110. - DOI - PMC - PubMed
-
- Karl D.L., Tupas L.R., Dore J., Christian J., Hebel D. The role of nitrogen fixation in biogeochemical cycling in the subtropical North Pacific Ocean. Nature. 1997;388:533–538. doi: 10.1038/41474. - DOI
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
