

Histone H3.3G34-Mutant Interneuron Progenitors Co-opt PDGFRA for Gliomagenesis
- PMID: 33259802
- PMCID: PMC7791404
- DOI: 10.1016/j.cell.2020.11.012
Histone H3.3G34-Mutant Interneuron Progenitors Co-opt PDGFRA for Gliomagenesis
Abstract
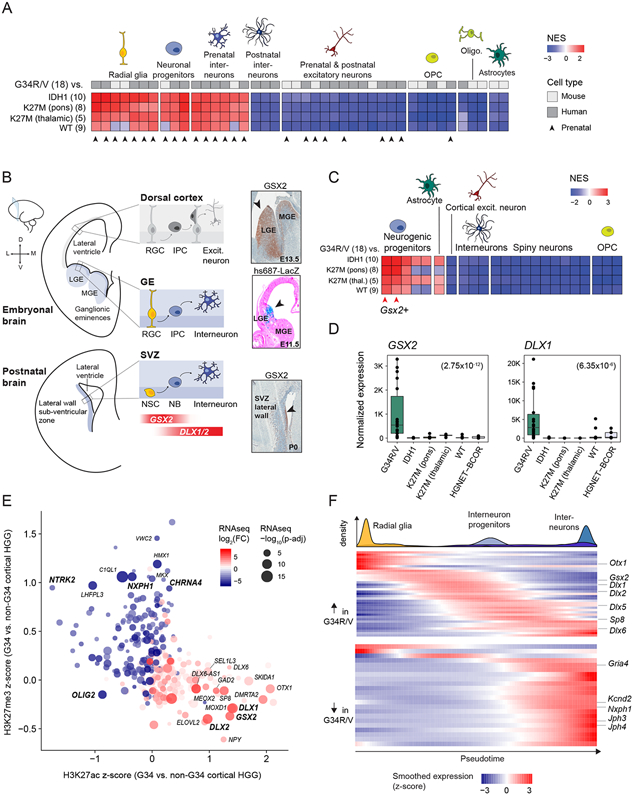
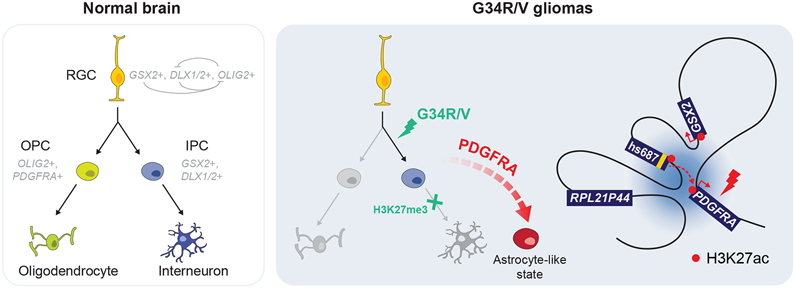
Histone H3.3 glycine 34 to arginine/valine (G34R/V) mutations drive deadly gliomas and show exquisite regional and temporal specificity, suggesting a developmental context permissive to their effects. Here we show that 50% of G34R/V tumors (n = 95) bear activating PDGFRA mutations that display strong selection pressure at recurrence. Although considered gliomas, G34R/V tumors actually arise in GSX2/DLX-expressing interneuron progenitors, where G34R/V mutations impair neuronal differentiation. The lineage of origin may facilitate PDGFRA co-option through a chromatin loop connecting PDGFRA to GSX2 regulatory elements, promoting PDGFRA overexpression and mutation. At the single-cell level, G34R/V tumors harbor dual neuronal/astroglial identity and lack oligodendroglial programs, actively repressed by GSX2/DLX-mediated cell fate specification. G34R/V may become dispensable for tumor maintenance, whereas mutant-PDGFRA is potently oncogenic. Collectively, our results open novel research avenues in deadly tumors. G34R/V gliomas are neuronal malignancies where interneuron progenitors are stalled in differentiation by G34R/V mutations and malignant gliogenesis is promoted by co-option of a potentially targetable pathway, PDGFRA signaling.
Keywords: GSX2; H3.3 G34R/V; PDGFRA; cell-of-origin; chromatin conformation; gliomas; interneuron progenitors; oncohistones; pediatric cancer; single-cell transcriptome.
Crown Copyright © 2020. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests P.B. and R.B. receive grant funding from the Novartis Institute of Biomedical Research for an unrelated project. J.R.H. has received compensation for consultation from Bayer for unrelated work.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
