

Shared Immunogenic Poly-Epitope Frameshift Mutations in Microsatellite Unstable Tumors
- PMID: 33259803
- PMCID: PMC8025604
- DOI: 10.1016/j.cell.2020.11.004
Shared Immunogenic Poly-Epitope Frameshift Mutations in Microsatellite Unstable Tumors
Abstract
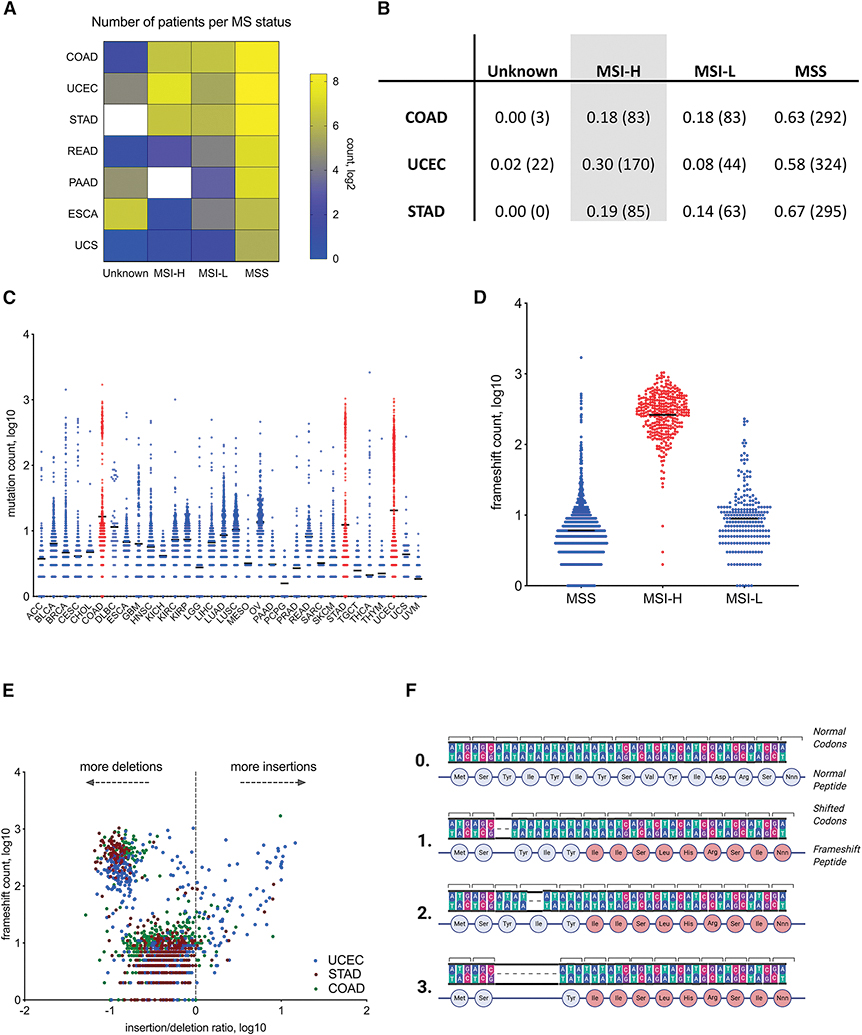
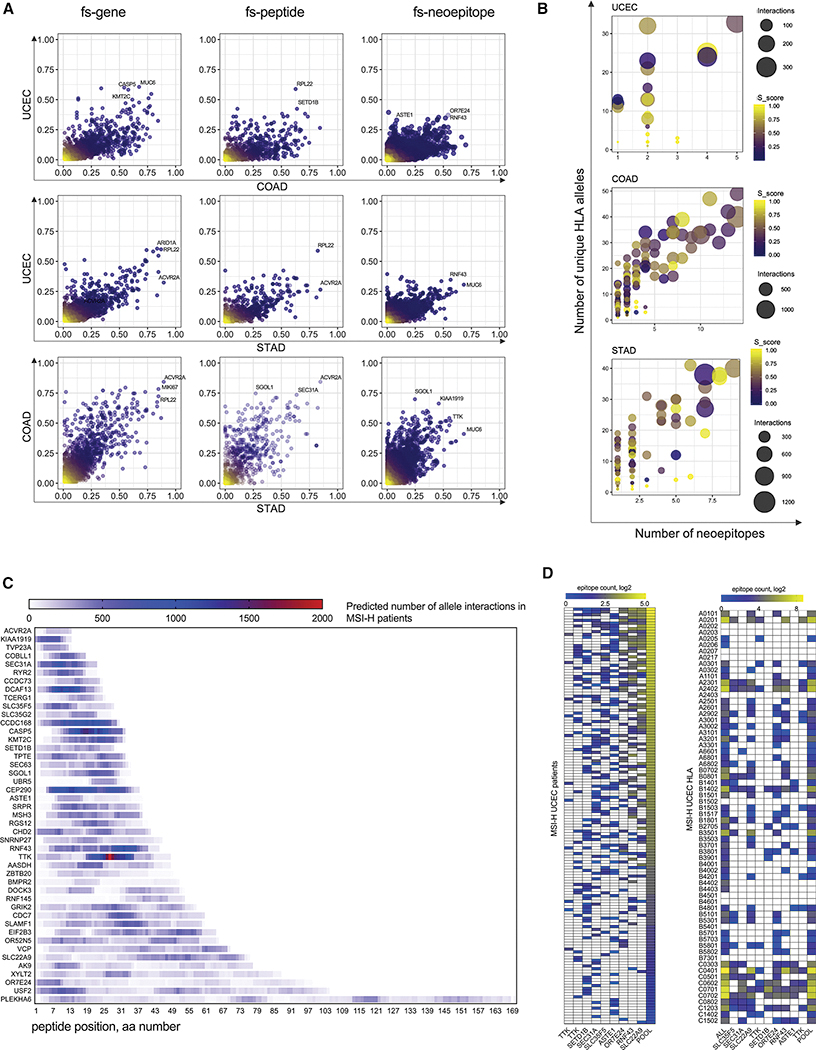
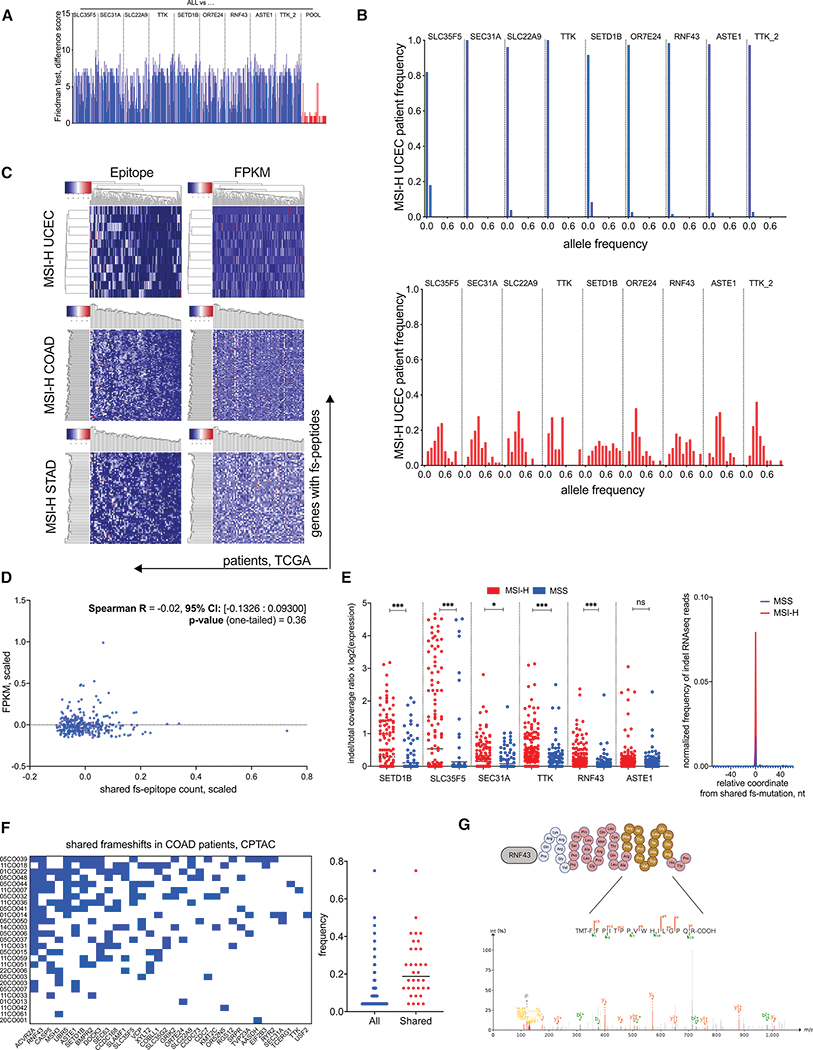
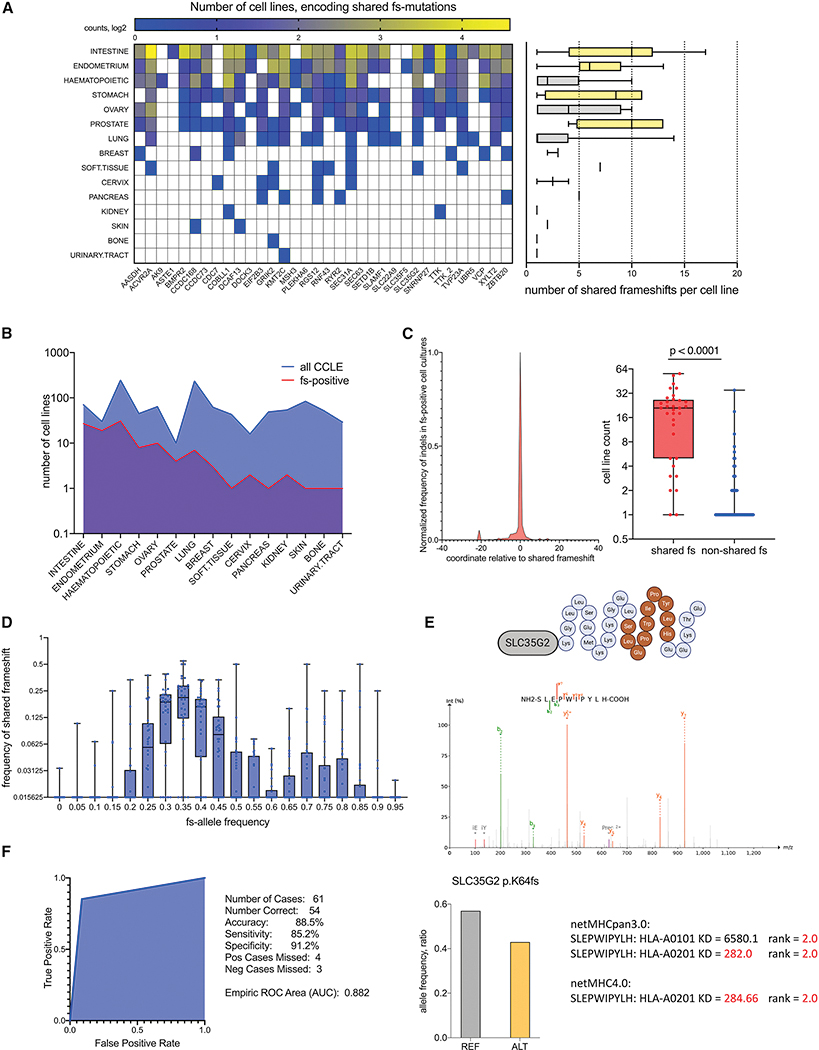
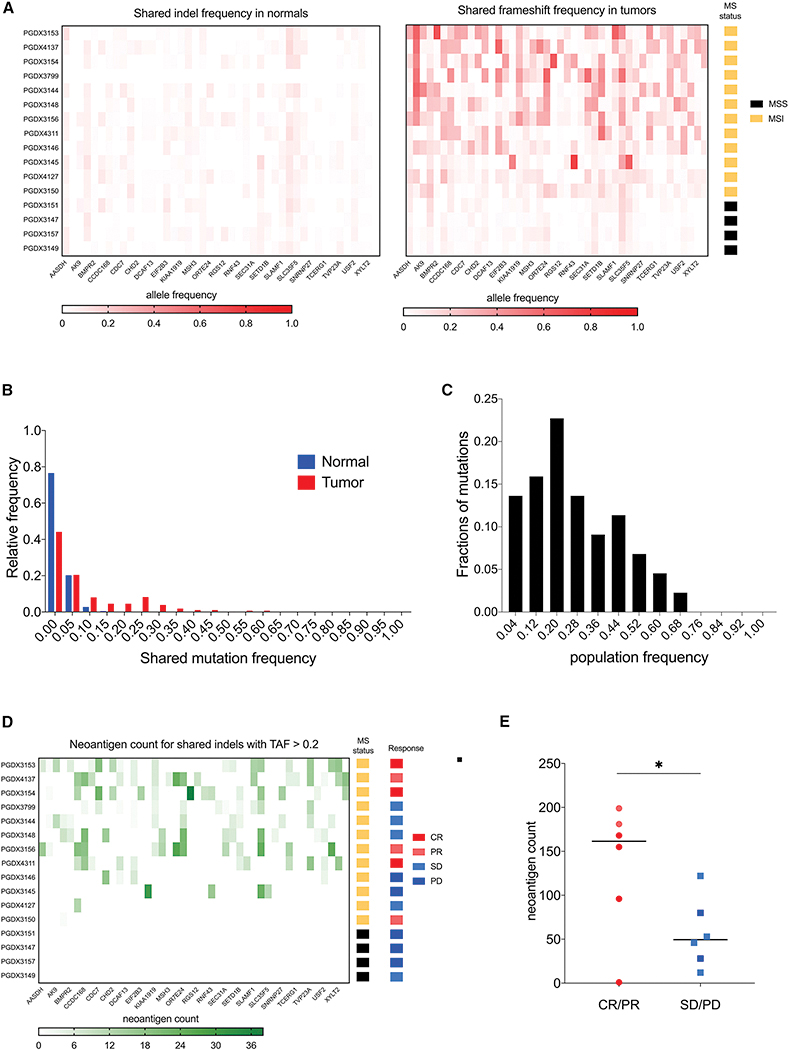
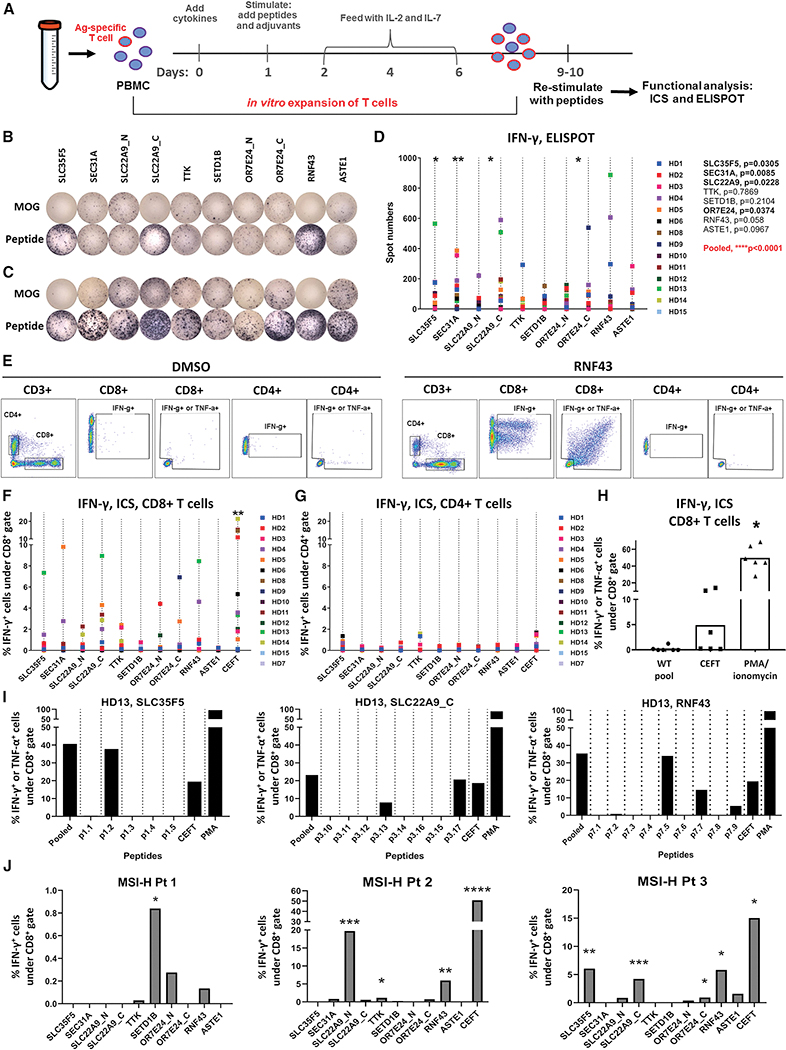
Microsatellite instability-high (MSI-H) tumors are characterized by high tumor mutation burden and responsiveness to checkpoint blockade. We identified tumor-specific frameshifts encoding multiple epitopes that originated from indel mutations shared among patients with MSI-H endometrial, colorectal, and stomach cancers. Epitopes derived from these shared frameshifts have high population occurrence rates, wide presence in many tumor subclones, and are predicted to bind to the most frequent MHC alleles in MSI-H patient cohorts. Neoantigens arising from these mutations are distinctly unlike self and viral antigens, signifying novel groups of potentially highly immunogenic tumor antigens. We further confirmed the immunogenicity of frameshift peptides in T cell stimulation experiments using blood mononuclear cells isolated from both healthy donors and MSI-H cancer patients. Our study uncovers the widespread occurrence and strong immunogenicity of tumor-specific antigens derived from shared frameshift mutations in MSI-H cancer and Lynch syndrome patients, suitable for the design of common "off-the-shelf" cancer vaccines.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests N.B. is an extramural member of the Parker Institute for Cancer Immunotherapy (PICI), receives research funds from Genentech, Oncovir, Regeneron and Dragonfly Therapeutics, and is on the advisory boards of Novartis, Roche, Avidea, Boehringer Ingelheim, Rome Therapeutics, BreakBio, Carisma Therapeutics, Roswell Park, and the Cancer Research Institute. C.B. is a PICI Bridge scholar. B.G. has received honoraria for speaking engagements from Merck, Bristol-Meyers Squibb, and Chugai Pharmaceuticals, has received research funding from Bristol-Meyers Squibb, and has been a compensated consultant for PMV Pharma and Rome Therapeutics of which he is a cofounder. V.R., C.C.B., N.B., B.G., S.B., and T.O. have a pending provisional patent application (no. 62/813,829 filed on March 5, 2019). The other authors have not declared any competing interests.
Figures
Comment in
-
Potential Use of Shared Frameshift Mutations in 'Off-the-Shelf' Neoantigen Vaccines.Trends Cancer. 2021 Mar;7(3):175-177. doi: 10.1016/j.trecan.2021.01.002. Epub 2021 Jan 23. Trends Cancer. 2021. PMID: 33500225
References
-
- Apcher S et al. (2011) ‘Major source of antigenic peptides for the MHC class I pathway is produced during the pioneer round of mRNA translation.’, Proceedings of the National Academy of Sciences of the United States of America, 108(28), pp. 11572–7. doi: 10.1073/pnas.1104104108. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
