

TYK2 Variants in B-Acute Lymphoblastic Leukaemia
- PMID: 33260630
- PMCID: PMC7761059
- DOI: 10.3390/genes11121434
TYK2 Variants in B-Acute Lymphoblastic Leukaemia
Abstract
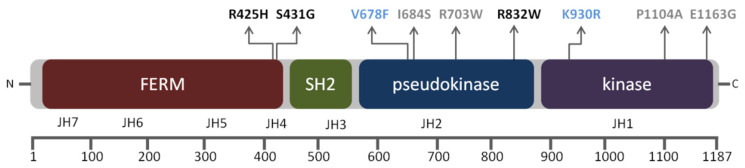
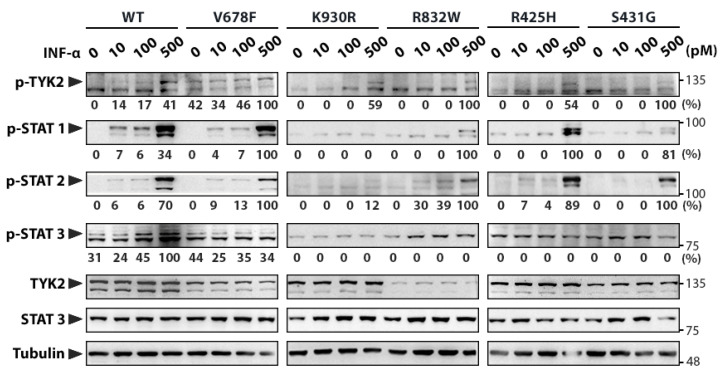
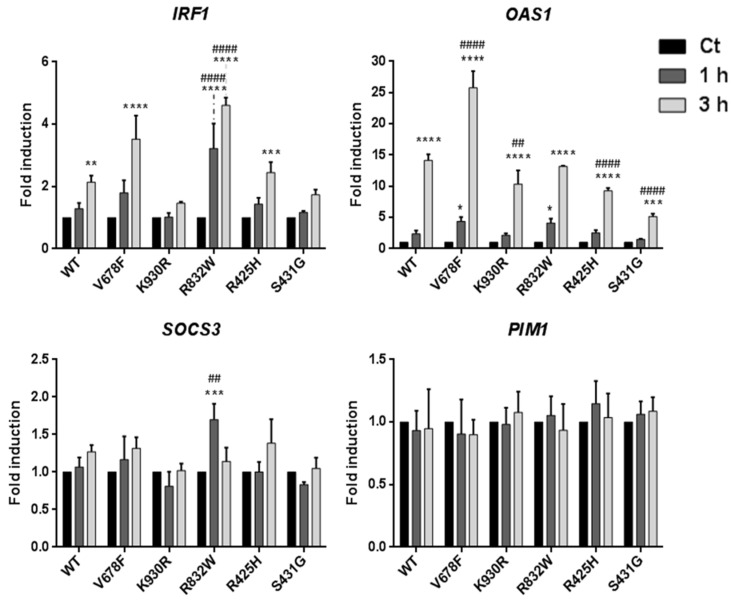
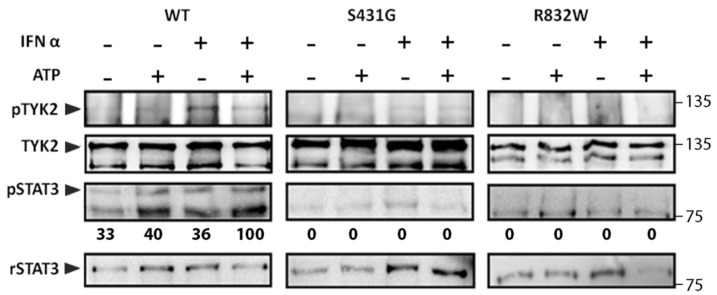
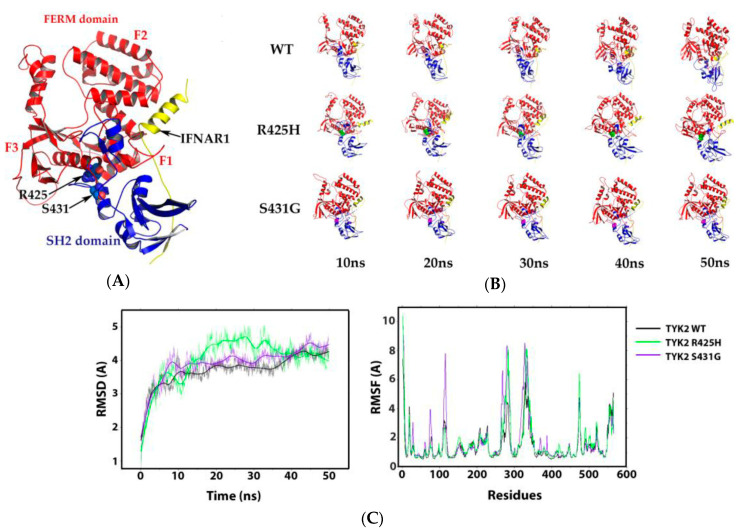
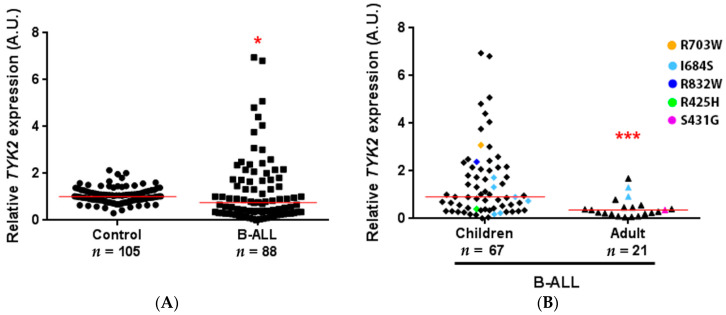
B-cell precursor acute lymphoblastic leukaemia (B-ALL) is a malignancy of lymphoid progenitor cells with altered genes including the Janus kinase (JAK) gene family. Among them, tyrosine kinase 2 (TYK2) is involved in signal transduction of cytokines such as interferon (IFN) α/β through IFN-α/β receptor alpha chain (IFNAR1). To search for disease-associated TYK2 variants, bone marrow samples from 62 B-ALL patients at diagnosis were analysed by next-generation sequencing. TYK2 variants were found in 16 patients (25.8%): one patient had a novel mutation at the four-point-one, ezrin, radixin, moesin (FERM) domain (S431G) and two patients had the rare variants rs150601734 or rs55882956 (R425H or R832W). To functionally characterise them, they were generated by direct mutagenesis, cloned in expression vectors, and transfected in TYK2-deficient cells. Under high-IFNα doses, the three variants were competent to phosphorylate STAT1/2. While R425H and R832W induced STAT1/2-target genes measured by qPCR, S431G behaved as the kinase-dead form of the protein. None of these variants phosphorylated STAT3 in in vitro kinase assays. Molecular dynamics simulation showed that TYK2/IFNAR1 interaction is not affected by these variants. Finally, qPCR analysis revealed diminished expression of TYK2 in B-ALL patients at diagnosis compared to that in healthy donors, further stressing the tumour immune surveillance role of TYK2.
Keywords: B-cell precursor acute lymphoblastic leukaemia; IFNα; IFNα/β receptor alpha chain (IFNAR1), next-generation sequencing; TYK2 expression; TYK2 variants; immune surveillance; molecular dynamics.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures
References
-
- Mullighan C.G., Zhang J., Harvey R.C., Collins-Underwood J.R., Schulman B.A., Phillips L.A., Tasian S.K., Loh M.L., Su X., Liu W., et al. JAK mutations in high-risk childhood acute lymphoblastic leukemia. Proc. Natl. Acad. Sci. USA. 2009;106:9414–9418. doi: 10.1073/pnas.0811761106. - DOI - PMC - PubMed
-
- Forero-Castro M., Robledo C., Benito R., Bodega-Mayor I., Rapado I., Hernandez-Sanchez M., Abaigar M., Maria Hernandez-Sanchez J., Quijada-Alamo M., Maria Sanchez-Pina J., et al. Mutations in TP53 and JAK2 are independent prognostic biomarkers in B-cell precursor acute lymphoblastic leukaemia. Br. J. Cancer. 2017;117:256–265. doi: 10.1038/bjc.2017.152. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
