

Regulation of murine copper homeostasis by members of the COMMD protein family
- PMID: 33262129
- PMCID: PMC7803461
- DOI: 10.1242/dmm.045963
Regulation of murine copper homeostasis by members of the COMMD protein family
Abstract
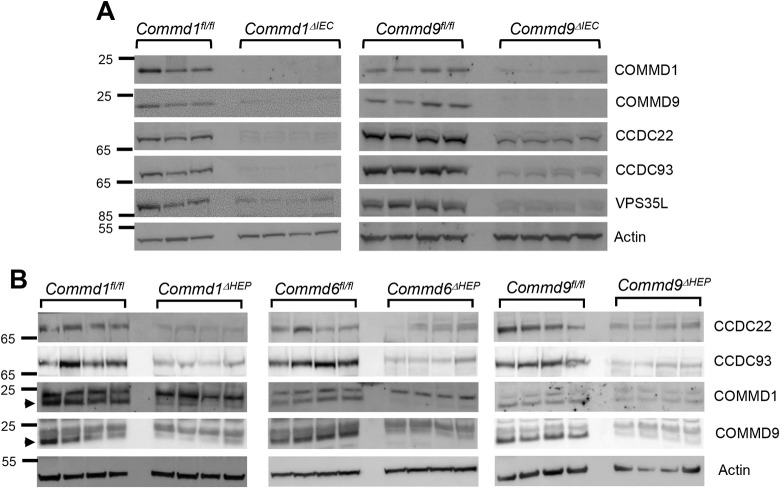
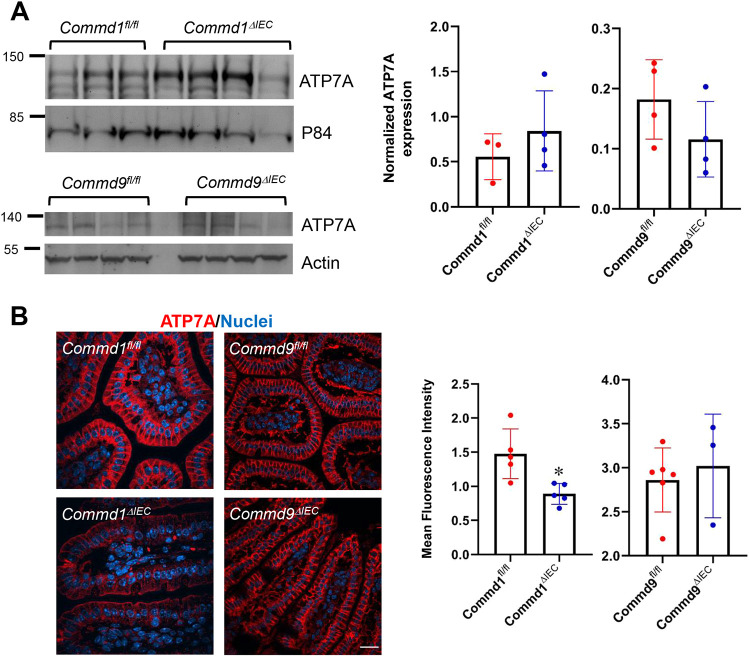
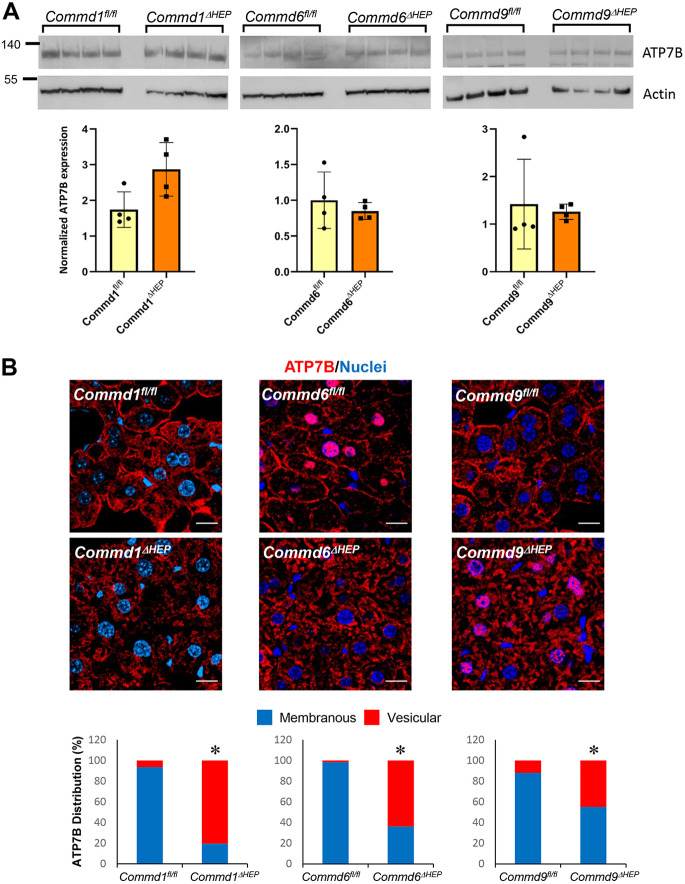
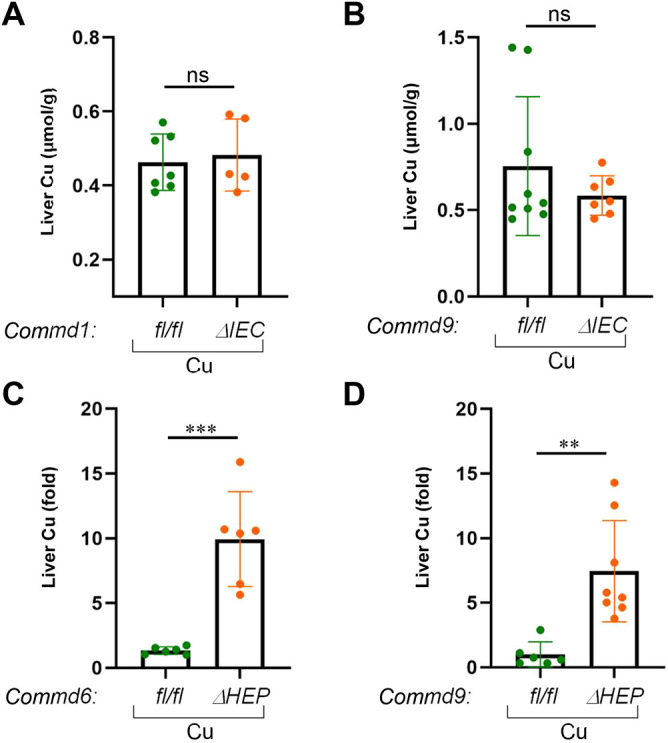
Copper is an essential transition metal for all eukaryotes. In mammals, intestinal copper absorption is mediated by the ATP7A copper transporter, whereas copper excretion occurs predominantly through the biliary route and is mediated by the paralog ATP7B. Both transporters have been shown to be recycled actively between the endosomal network and the plasma membrane by a molecular machinery known as the COMMD/CCDC22/CCDC93 or CCC complex. In fact, mutations in COMMD1 can lead to impaired biliary copper excretion and liver pathology in dogs and in mice with liver-specific Commd1 deficiency, recapitulating aspects of this phenotype. Nonetheless, the role of the CCC complex in intestinal copper absorption in vivo has not been studied, and the potential redundancy of various COMMD family members has not been tested. In this study, we examined copper homeostasis in enterocyte-specific and hepatocyte-specific COMMD gene-deficient mice. We found that, in contrast to effects in cell lines in culture, COMMD protein deficiency induced minimal changes in ATP7A in enterocytes and did not lead to altered copper levels under low- or high-copper diets, suggesting that regulation of ATP7A in enterocytes is not of physiological consequence. By contrast, deficiency of any of three COMMD genes (Commd1, Commd6 or Commd9) resulted in hepatic copper accumulation under high-copper diets. We found that each of these deficiencies caused destabilization of the entire CCC complex and suggest that this might explain their shared phenotype. Overall, we conclude that the CCC complex plays an important role in ATP7B endosomal recycling and function.
Keywords: ATP7A; ATP7B; COMMD proteins; Copper homeostasis; Copper transporters; Endosomal trafficking.
© 2021. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures
References
-
- Bartuzi P., Billadeau D. D., Favier R., Rong S., Dekker D., Fedoseienko A., Fieten H., Wijers M., Levels J. H., Huijkman N. et al. (2016). CCC- and WASH-mediated endosomal sorting of LDLR is required for normal clearance of circulating LDL. Nat. Commun. 7, 10961 10.1038/ncomms10961 - DOI - PMC - PubMed
-
- Burstein E., Hoberg J. E., Wilkinson A. S., Rumble J. M., Csomos R. A., Komarck C. M., Maine G. N., Wilkinson J. C., Mayo M. W. and Duckett C. S. (2005). COMMD proteins: a novel family of structural and functional homologs of MURR1. J. Biol. Chem. 280, 22222-22232. 10.1074/jbc.M501928200 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
