

High-Frequency Neuronal Bursting is Essential for Circadian and Sleep Behaviors in Drosophila
- PMID: 33262246
- PMCID: PMC7842748
- DOI: 10.1523/JNEUROSCI.2322-20.2020
High-Frequency Neuronal Bursting is Essential for Circadian and Sleep Behaviors in Drosophila
Abstract
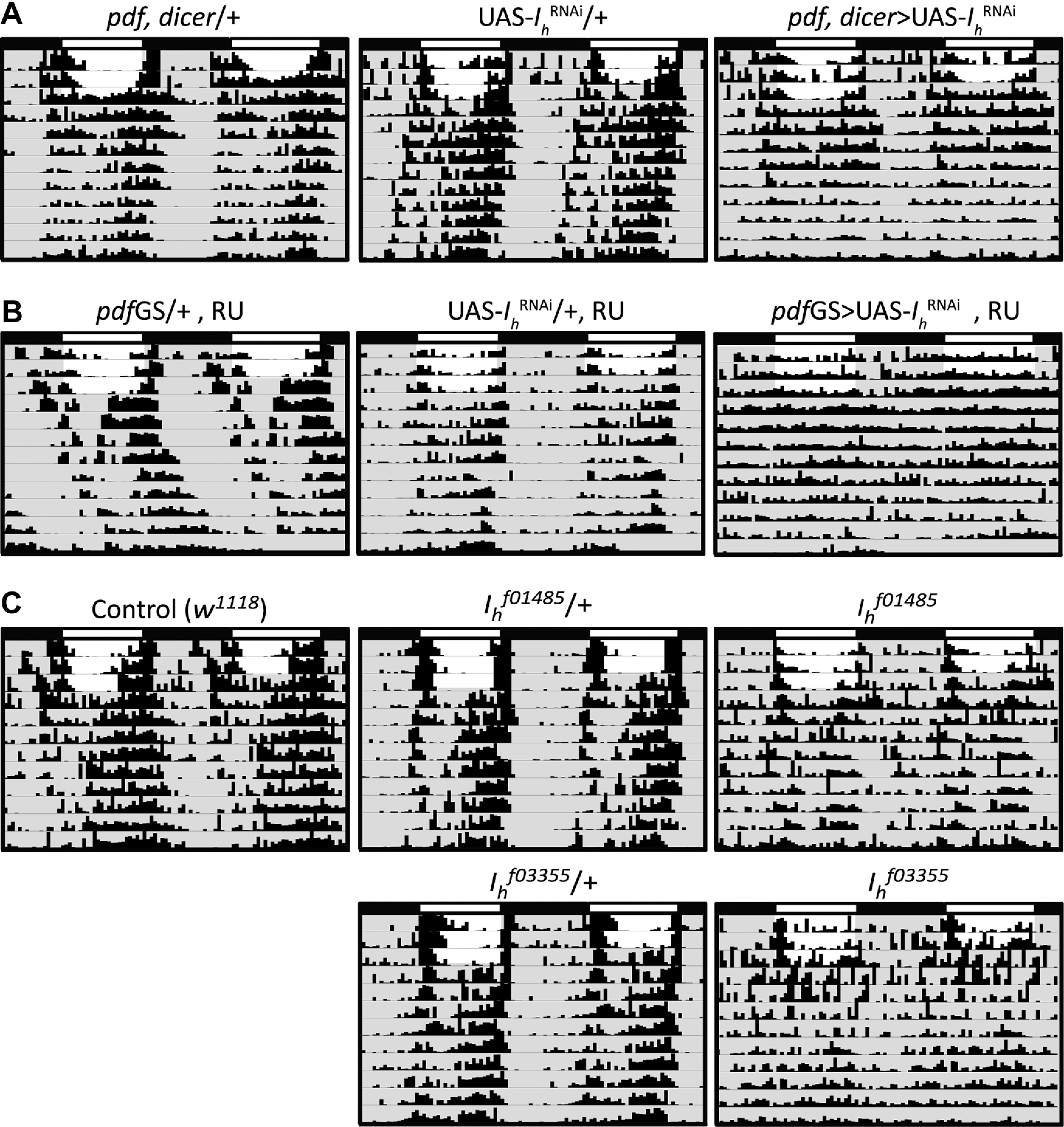
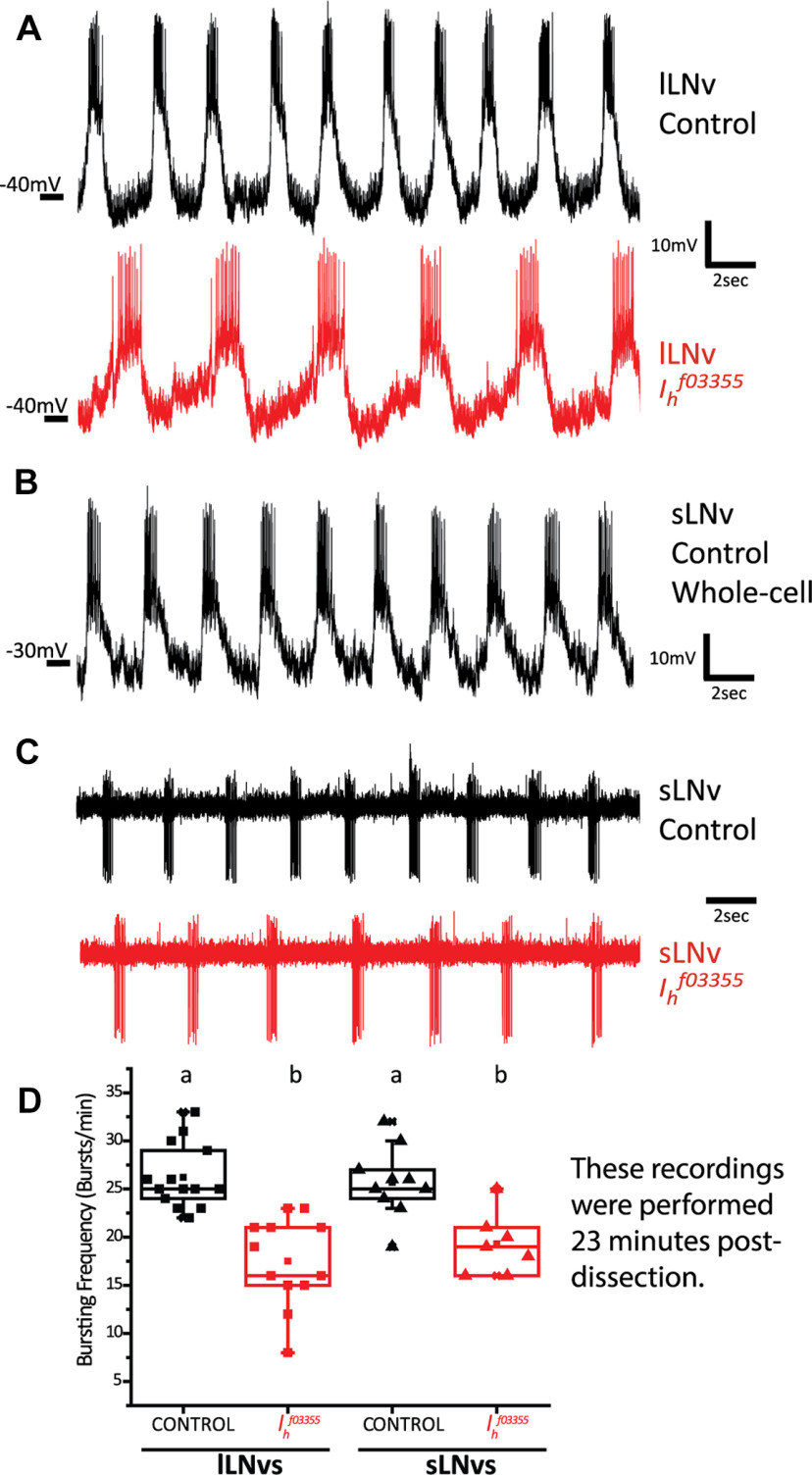
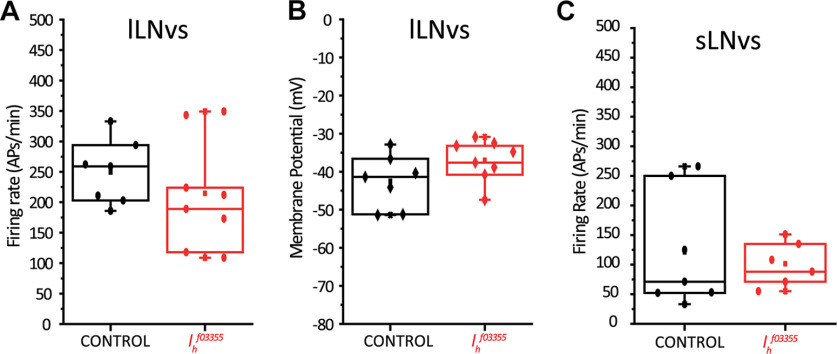
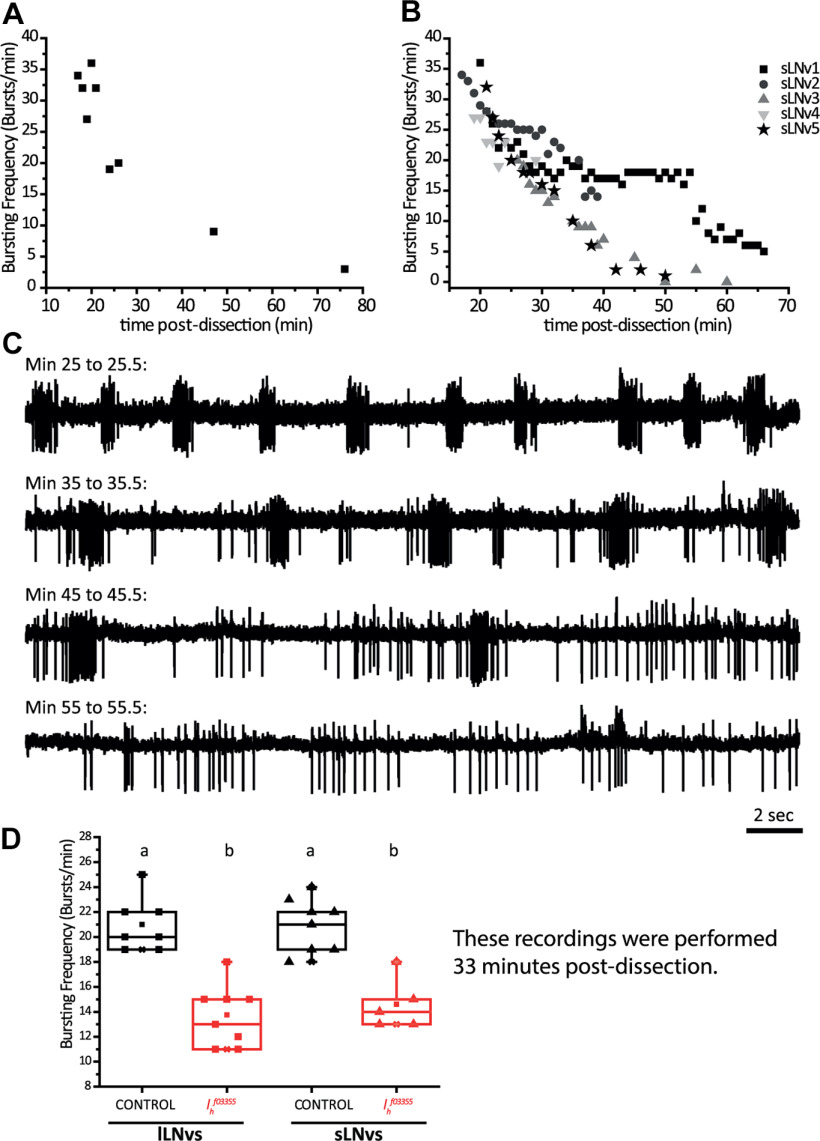
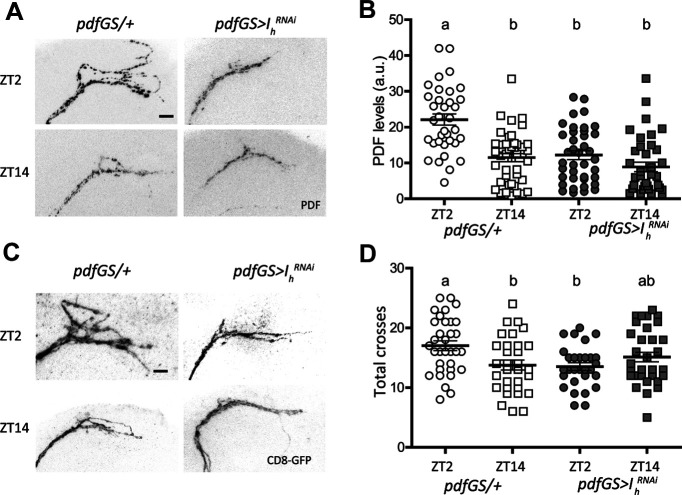
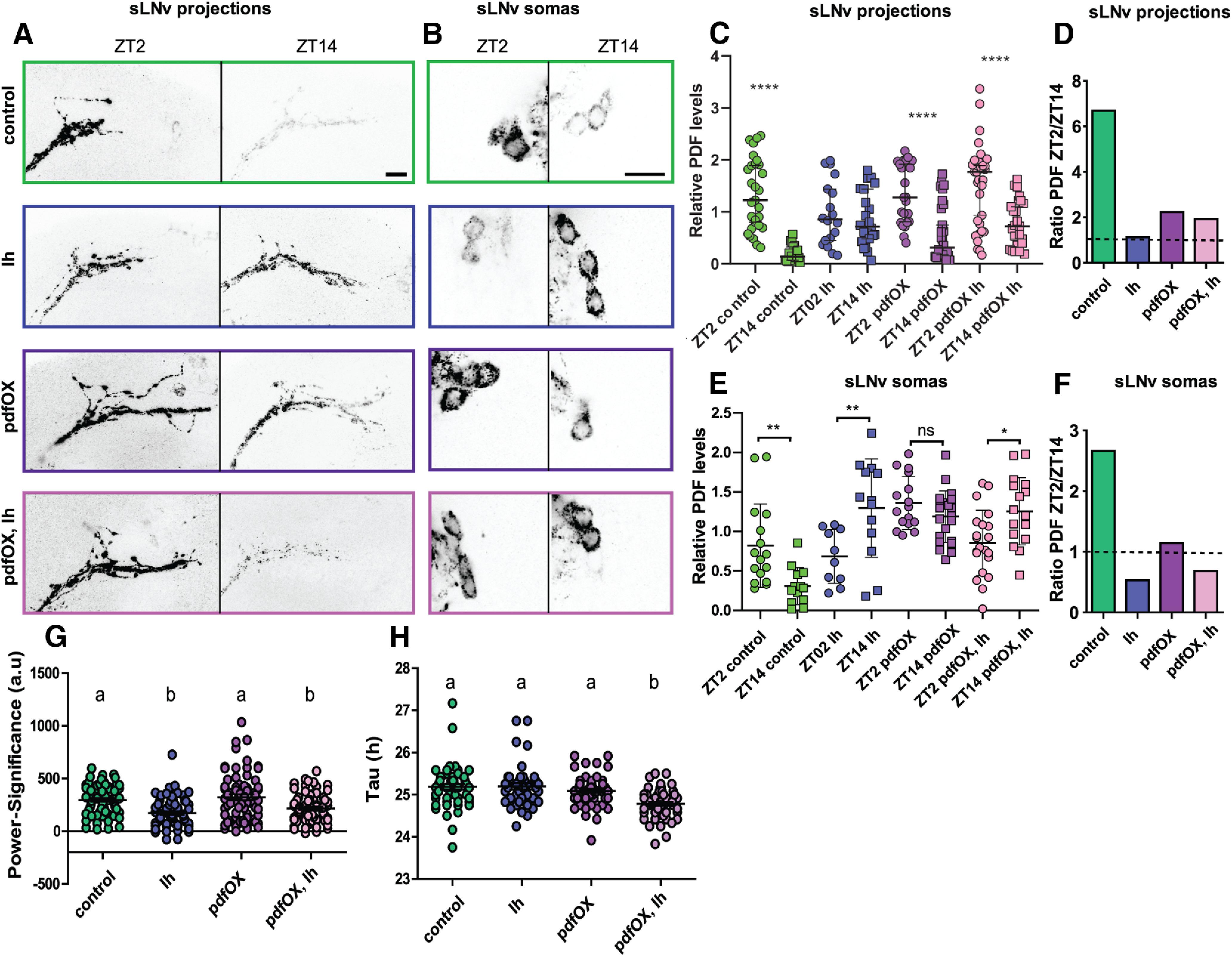
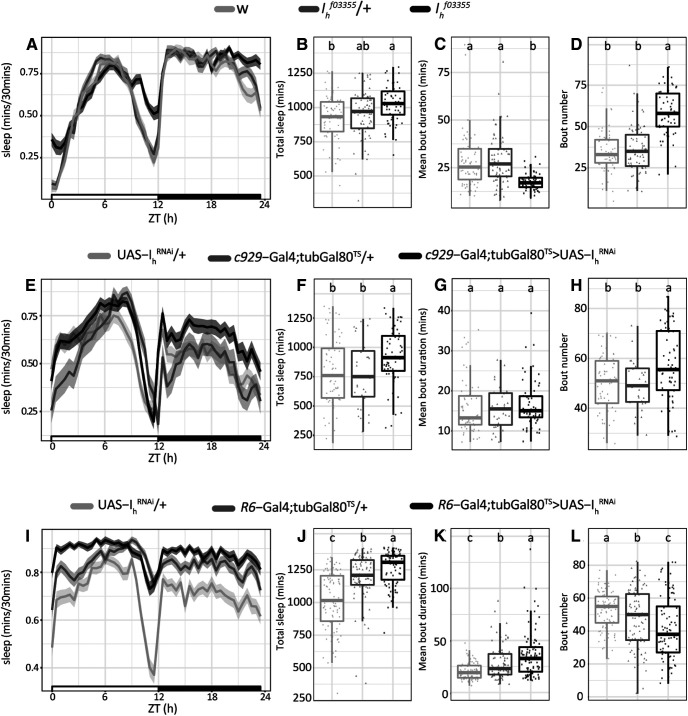
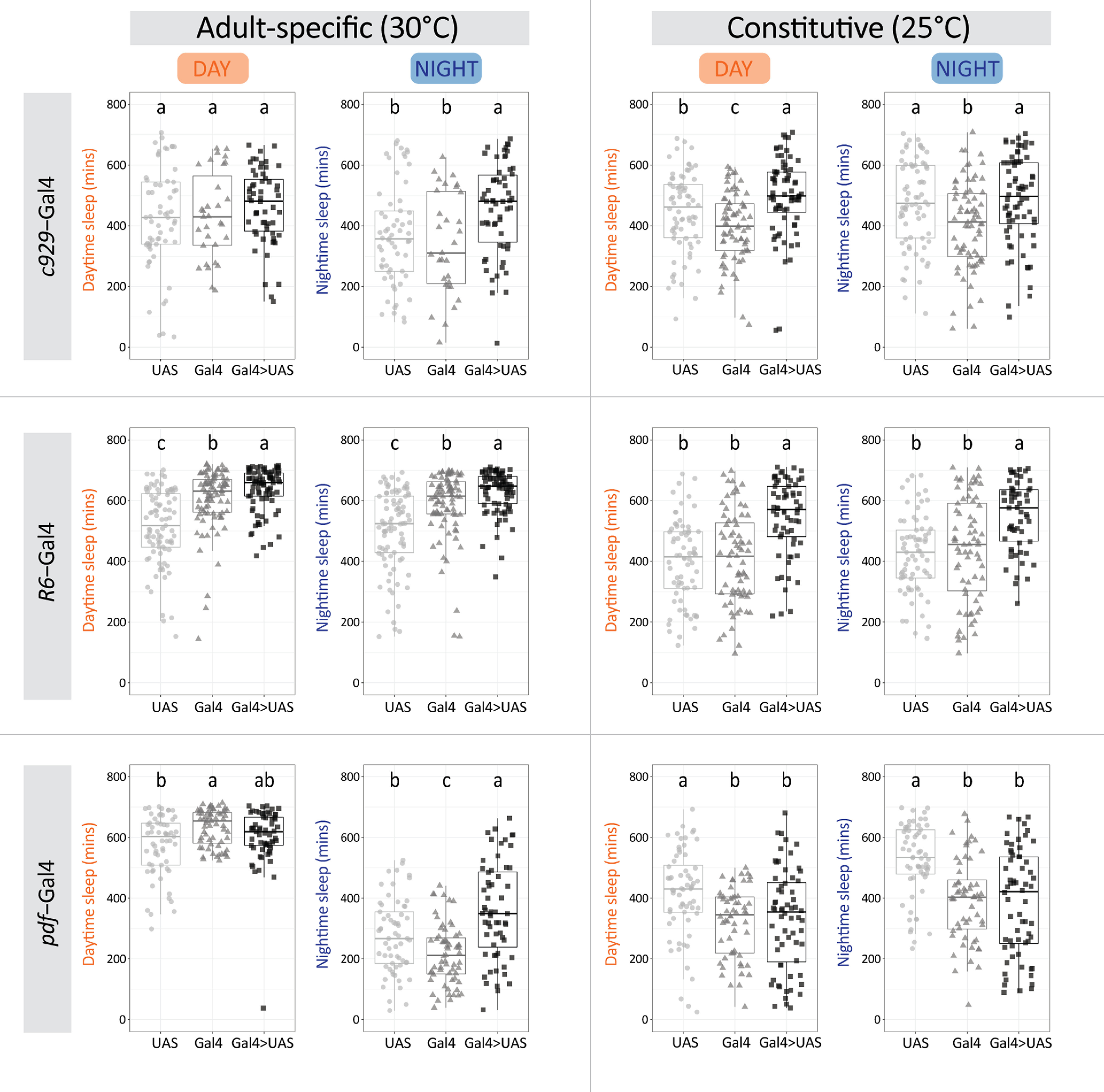
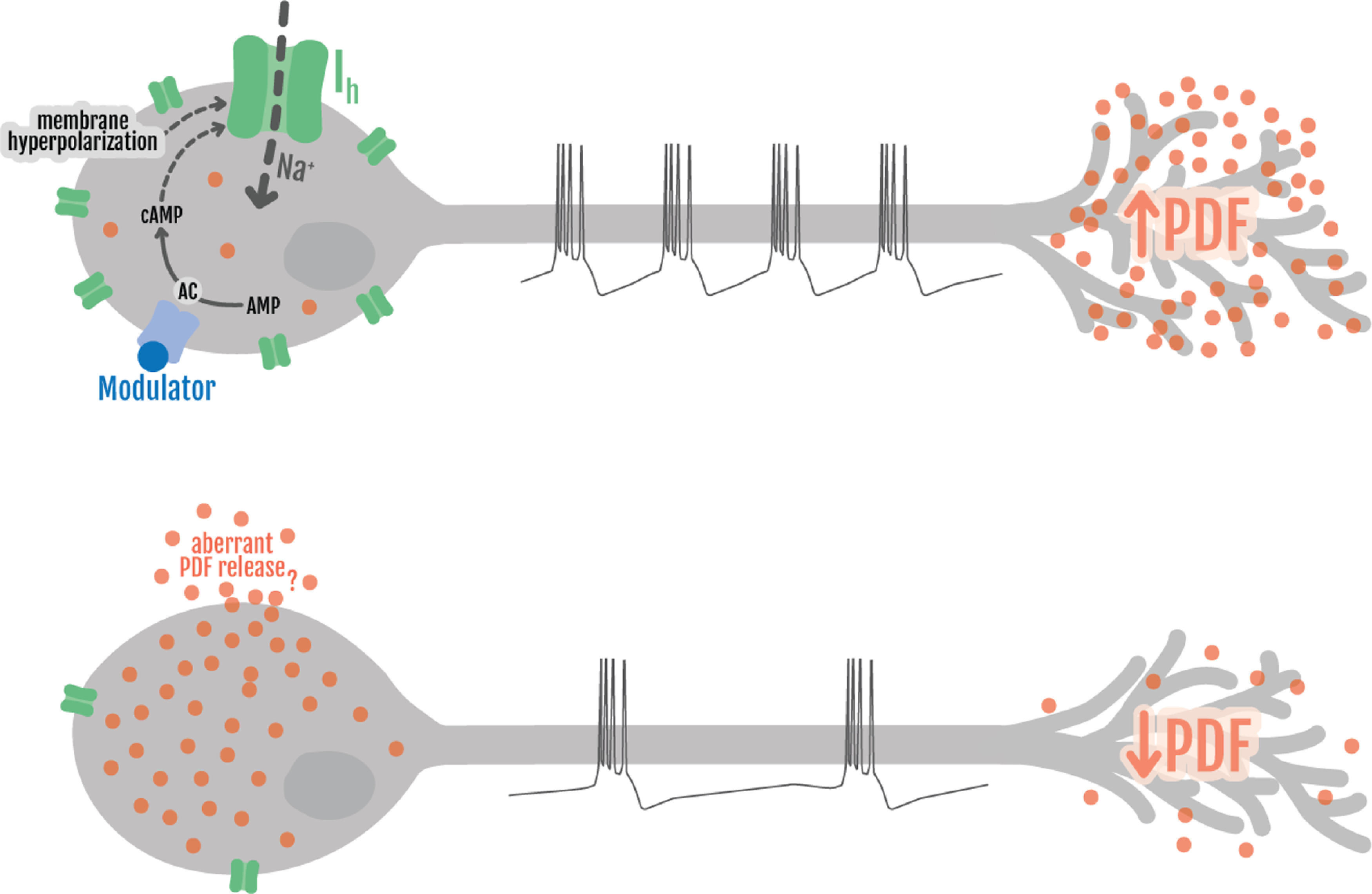
Circadian rhythms have been extensively studied in Drosophila; however, still little is known about how the electrical properties of clock neurons are specified. We have performed a behavioral genetic screen through the downregulation of candidate ion channels in the lateral ventral neurons (LNvs) and show that the hyperpolarization-activated cation current Ih is important for the behaviors that the LNvs influence: temporal organization of locomotor activity, analyzed in males, and sleep, analyzed in females. Using whole-cell patch clamp electrophysiology we demonstrate that small LNvs (sLNvs) are bursting neurons, and that Ih is necessary to achieve the high-frequency bursting firing pattern characteristic of both types of LNvs in females. Since firing in bursts has been associated to neuropeptide release, we hypothesized that Ih would be important for LNvs communication. Indeed, herein we demonstrate that Ih is fundamental for the recruitment of pigment dispersing factor (PDF) filled dense core vesicles (DCVs) to the terminals at the dorsal protocerebrum and for their timed release, and hence for the temporal coordination of circadian behaviors.SIGNIFICANCE STATEMENT Ion channels are transmembrane proteins with selective permeability to specific charged particles. The rich repertoire of parameters that may gate their opening state, such as voltage-sensitivity, modulation by second messengers and specific kinetics, make this protein family a determinant of neuronal identity. Ion channel structure is evolutionary conserved between vertebrates and invertebrates, making any discovery easily translatable. Through a screen to uncover ion channels with roles in circadian rhythms, we have identified the Ih channel as an important player in a subset of clock neurons of the fruit fly. We show that lateral ventral neurons (LNvs) need Ih to fire action potentials in a high-frequency bursting mode and that this is important for peptide transport and the control of behavior.
Keywords: Drosophila melanogaster; HCN; Ih; bursting neuron; ion channel; pigment dispersing factor.
Copyright © 2021 the authors.
Figures
Similar articles
-
Electrical hyperexcitation of lateral ventral pacemaker neurons desynchronizes downstream circadian oscillators in the fly circadian circuit and induces multiple behavioral periods.J Neurosci. 2006 Jan 11;26(2):479-89. doi: 10.1523/JNEUROSCI.3915-05.2006. J Neurosci. 2006. PMID: 16407545 Free PMC article.
-
Alzheimer's disease-associated tau alters Drosophila circadian activity, sleep and clock neuron electrophysiology.Neurobiol Dis. 2019 Oct;130:104507. doi: 10.1016/j.nbd.2019.104507. Epub 2019 Jun 14. Neurobiol Dis. 2019. PMID: 31207389
-
Coupling Neuropeptide Levels to Structural Plasticity in Drosophila Clock Neurons.Curr Biol. 2020 Aug 17;30(16):3154-3166.e4. doi: 10.1016/j.cub.2020.06.009. Epub 2020 Jul 2. Curr Biol. 2020. PMID: 32619484
-
Circadian Rhythms and Sleep in Drosophila melanogaster.Genetics. 2017 Apr;205(4):1373-1397. doi: 10.1534/genetics.115.185157. Genetics. 2017. PMID: 28360128 Free PMC article. Review.
-
Mechanisms of clock output in the Drosophila circadian pacemaker system.J Biol Rhythms. 2006 Dec;21(6):445-57. doi: 10.1177/0748730406293910. J Biol Rhythms. 2006. PMID: 17107935 Review.
Cited by
-
Decapentaplegic Acutely Defines the Connectivity of Central Pacemaker Neurons in Drosophila.J Neurosci. 2021 Oct 6;41(40):8338-8350. doi: 10.1523/JNEUROSCI.0397-21.2021. Epub 2021 Aug 24. J Neurosci. 2021. PMID: 34429376 Free PMC article.
-
A unified framework to model synaptic dynamics during the sleep-wake cycle.PLoS Biol. 2025 Jun 12;23(6):e3003198. doi: 10.1371/journal.pbio.3003198. eCollection 2025 Jun. PLoS Biol. 2025. PMID: 40504782 Free PMC article.
-
Temperature-dependent modulation of light-induced circadian responses in Drosophila melanogaster.EMBO J. 2025 Aug;44(16):4552-4576. doi: 10.1038/s44318-025-00499-w. Epub 2025 Jun 30. EMBO J. 2025. PMID: 40588550 Free PMC article.
-
Circadian Rhythm Regulation by Pacemaker Neuron Chloride Oscillation in Flies.Physiology (Bethesda). 2024 May 1;39(3):0. doi: 10.1152/physiol.00006.2024. Epub 2024 Feb 27. Physiology (Bethesda). 2024. PMID: 38411570 Free PMC article. Review.
-
An evolutionarily conserved cation channel tunes the sensitivity of gustatory neurons to ephaptic inhibition in Drosophila.Proc Natl Acad Sci U S A. 2025 Jan 21;122(3):e2413134122. doi: 10.1073/pnas.2413134122. Epub 2025 Jan 17. Proc Natl Acad Sci U S A. 2025. PMID: 39823301 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials