

The Functional Impact of Mitochondrial Structure Across Subcellular Scales
- PMID: 33262702
- PMCID: PMC7686514
- DOI: 10.3389/fphys.2020.541040
The Functional Impact of Mitochondrial Structure Across Subcellular Scales
Abstract
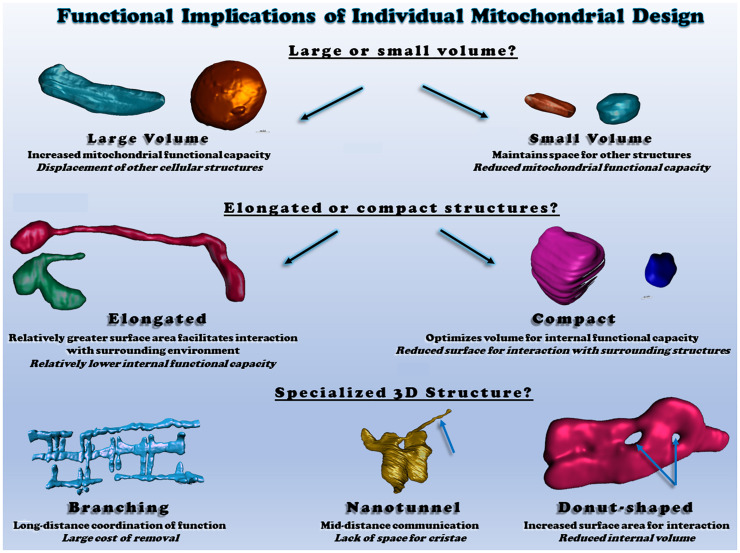
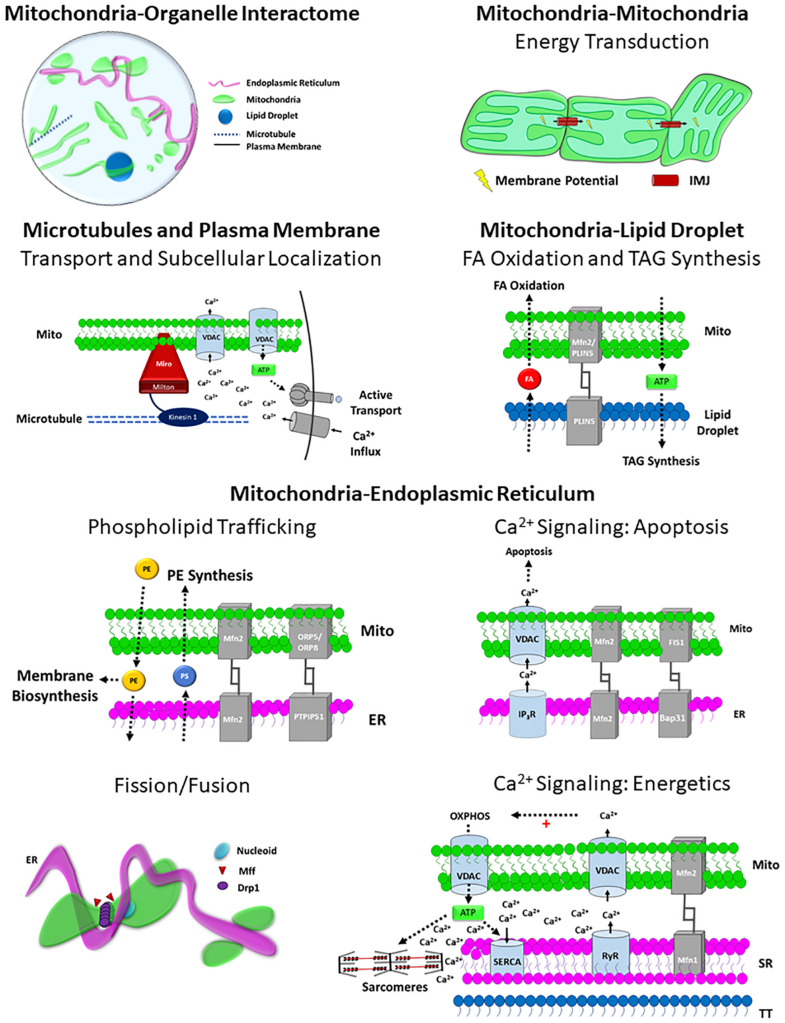
Mitochondria are key determinants of cellular health. However, the functional role of mitochondria varies from cell to cell depending on the relative demands for energy distribution, metabolite biosynthesis, and/or signaling. In order to support the specific needs of different cell types, mitochondrial functional capacity can be optimized in part by modulating mitochondrial structure across several different spatial scales. Here we discuss the functional implications of altering mitochondrial structure with an emphasis on the physiological trade-offs associated with different mitochondrial configurations. Within a mitochondrion, increasing the amount of cristae in the inner membrane improves capacity for energy conversion and free radical-mediated signaling but may come at the expense of matrix space where enzymes critical for metabolite biosynthesis and signaling reside. Electrically isolating individual cristae could provide a protective mechanism to limit the spread of dysfunction within a mitochondrion but may also slow the response time to an increase in cellular energy demand. For individual mitochondria, those with relatively greater surface areas can facilitate interactions with the cytosol or other organelles but may be more costly to remove through mitophagy due to the need for larger phagophore membranes. At the network scale, a large, stable mitochondrial reticulum can provide a structural pathway for energy distribution and communication across long distances yet also enable rapid spreading of localized dysfunction. Highly dynamic mitochondrial networks allow for frequent content mixing and communication but require constant cellular remodeling to accommodate the movement of mitochondria. The formation of contact sites between mitochondria and several other organelles provides a mechanism for specialized communication and direct content transfer between organelles. However, increasing the number of contact sites between mitochondria and any given organelle reduces the mitochondrial surface area available for contact sites with other organelles as well as for metabolite exchange with cytosol. Though the precise mechanisms guiding the coordinated multi-scale mitochondrial configurations observed in different cell types have yet to be elucidated, it is clear that mitochondrial structure is tailored at every level to optimize mitochondrial function to meet specific cellular demands.
Keywords: cristae; energetics; mitochondria; mitochondrial dynamics; mitochondrial networks; organelle interaction.
Copyright © 2020 Glancy, Kim, Katti and Willingham.
Figures


References
Publication types
LinkOut - more resources
Full Text Sources