

Central processing of leg proprioception in Drosophila
- PMID: 33263281
- PMCID: PMC7752136
- DOI: 10.7554/eLife.60299
Central processing of leg proprioception in Drosophila
Abstract
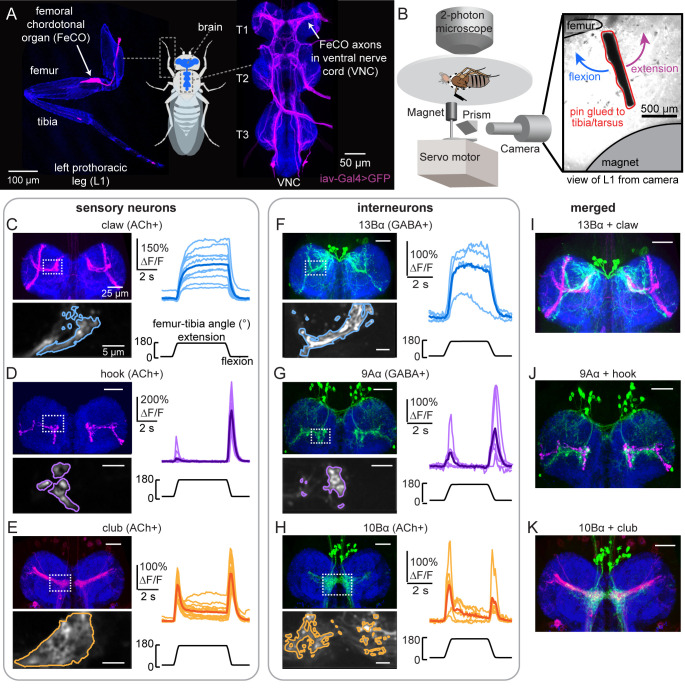
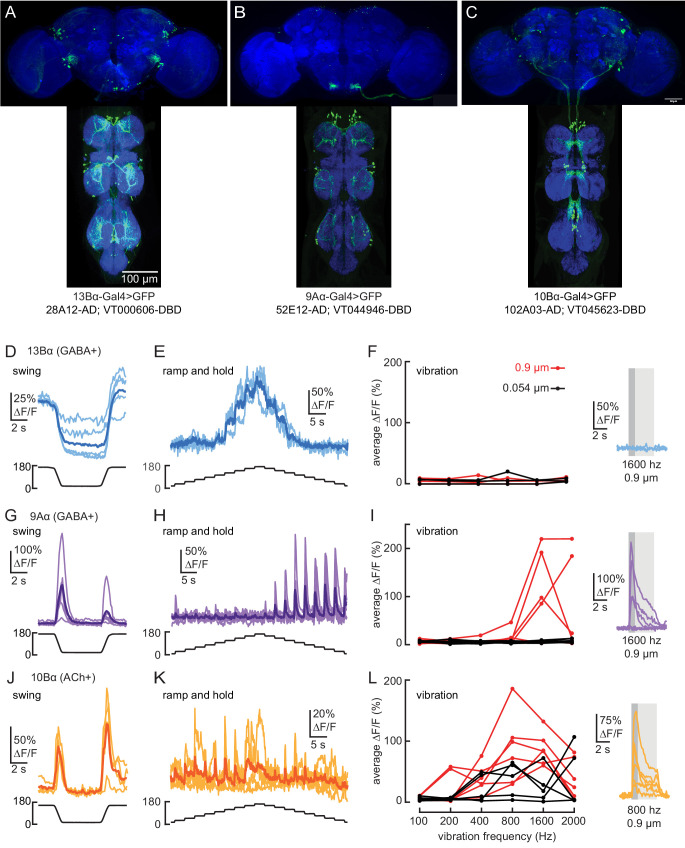
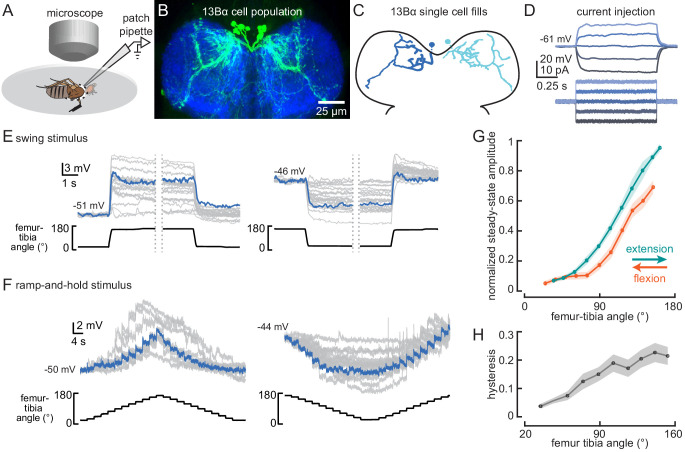
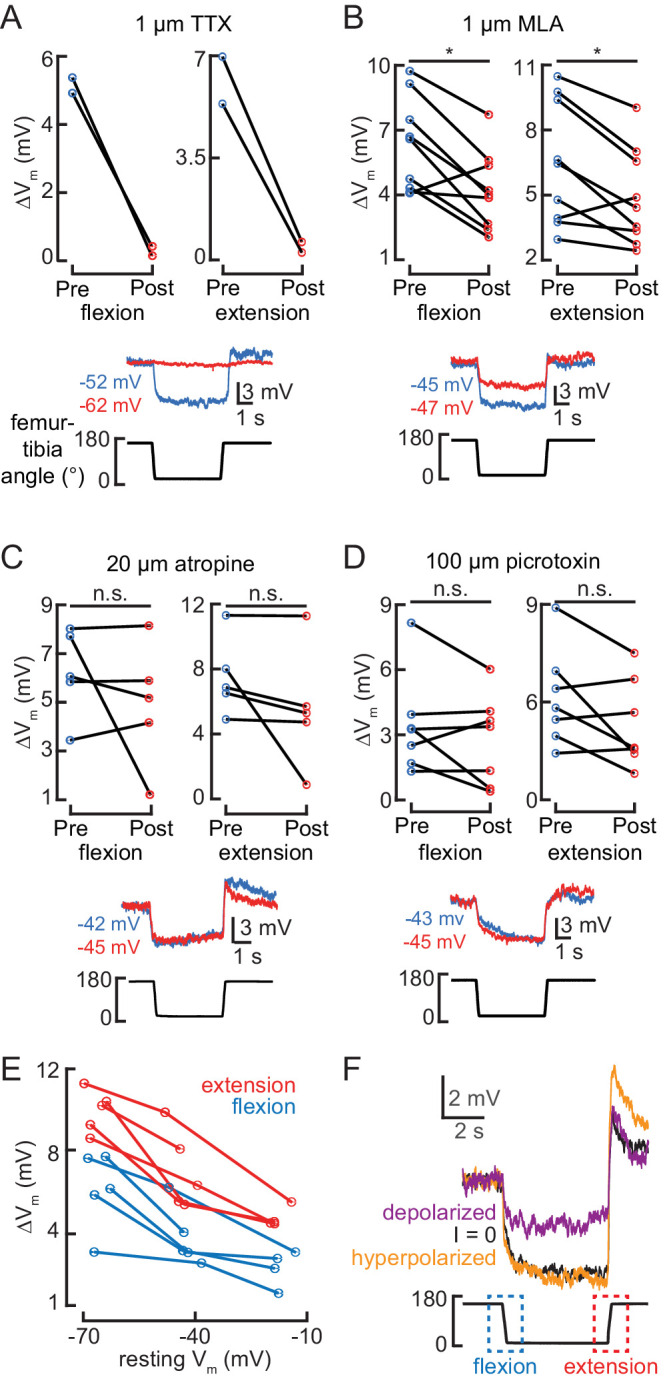
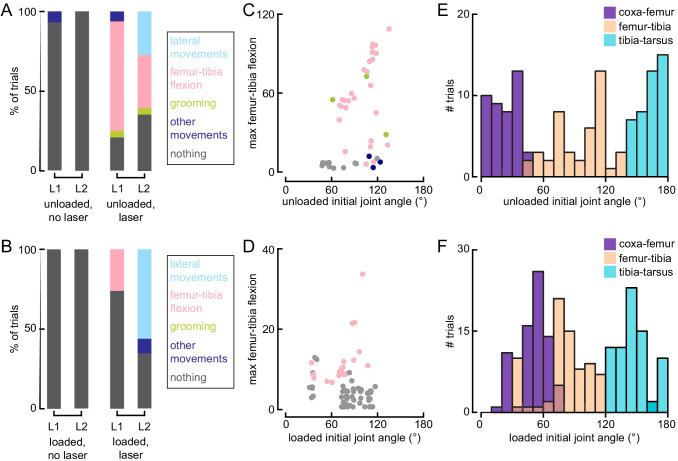
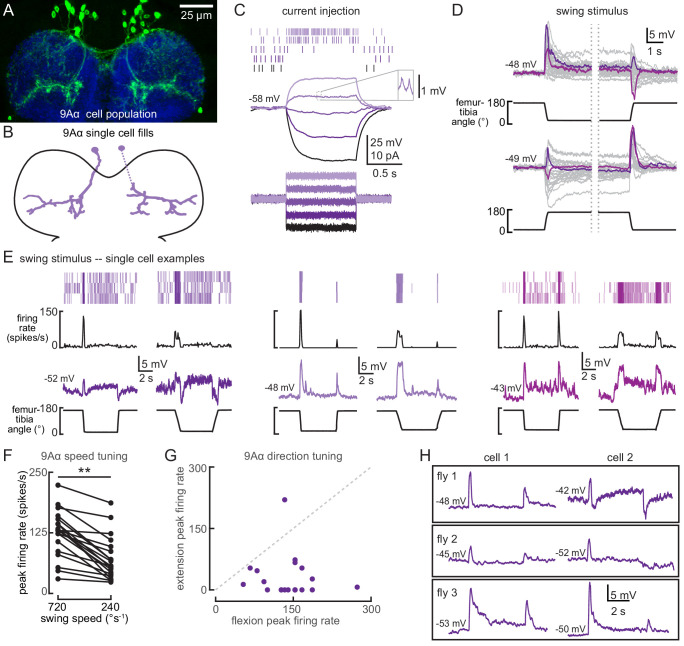
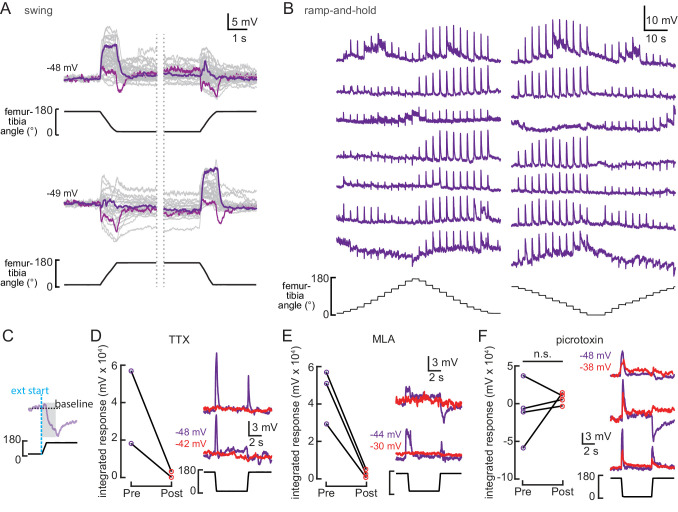
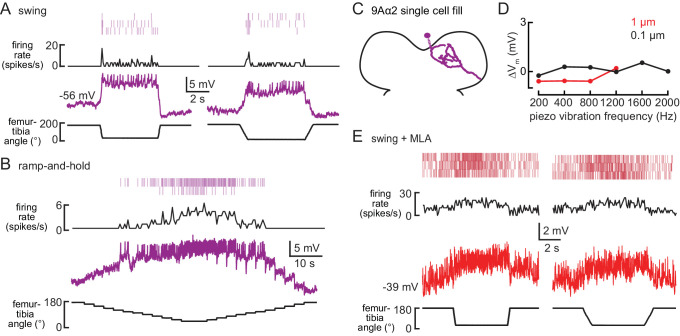
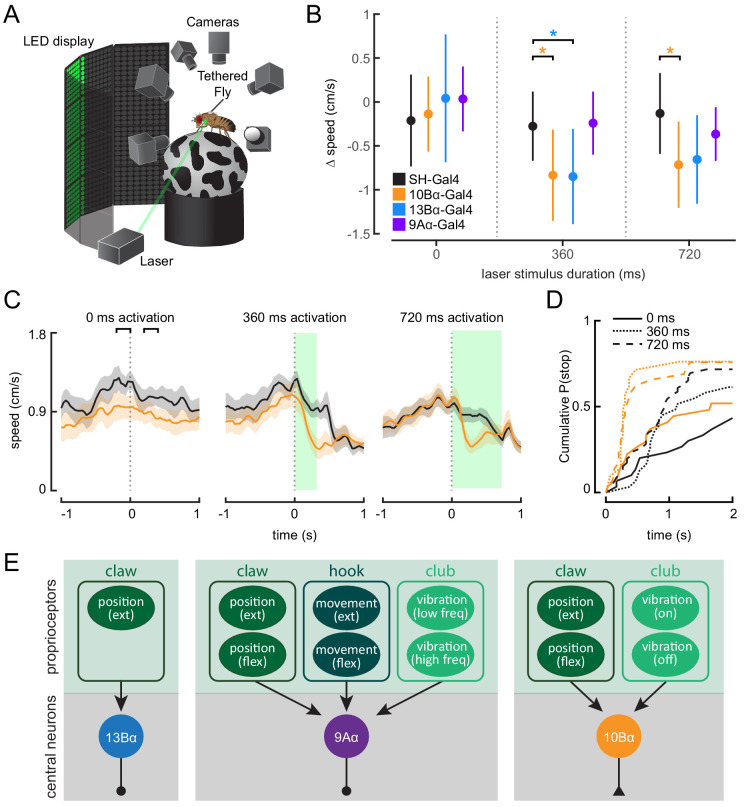
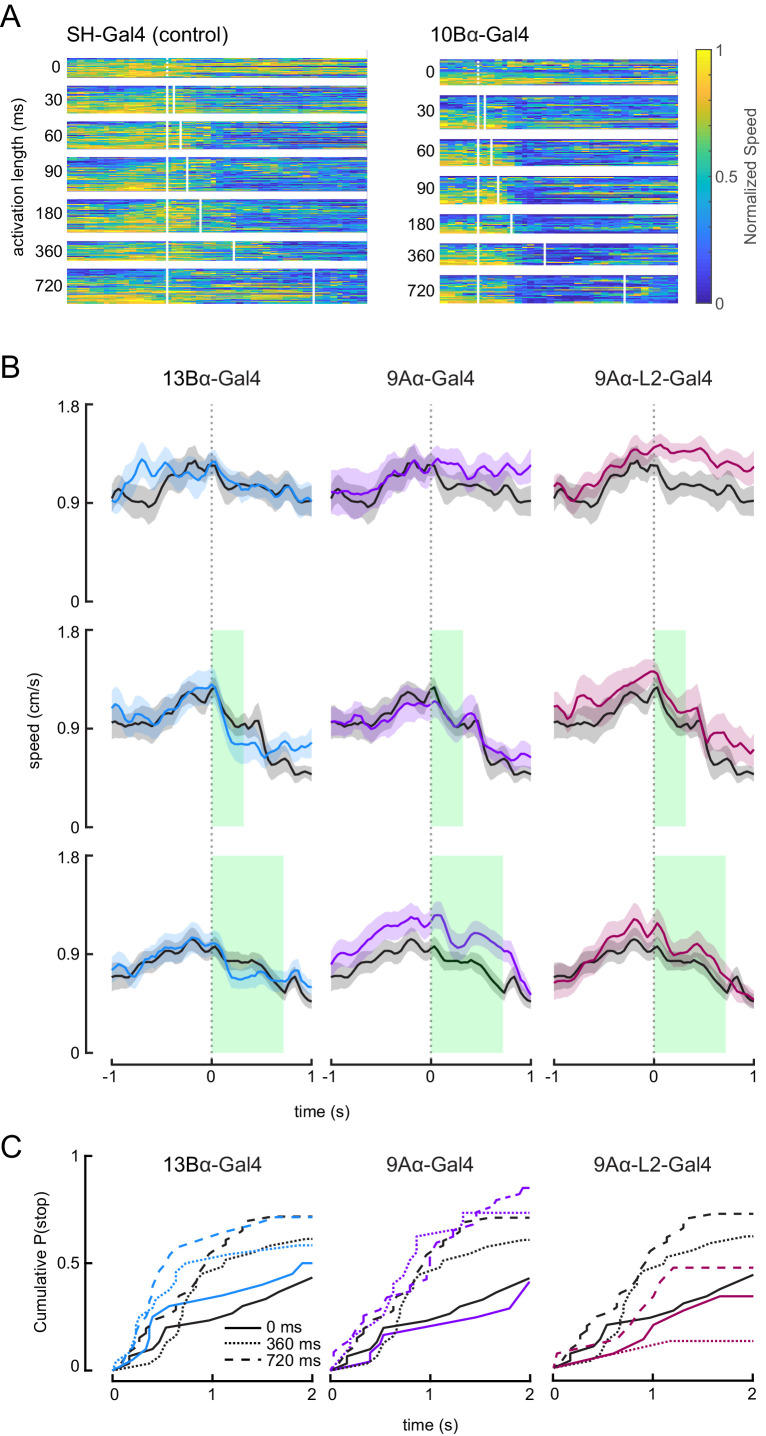
Proprioception, the sense of self-movement and position, is mediated by mechanosensory neurons that detect diverse features of body kinematics. Although proprioceptive feedback is crucial for accurate motor control, little is known about how downstream circuits transform limb sensory information to guide motor output. Here we investigate neural circuits in Drosophila that process proprioceptive information from the fly leg. We identify three cell types from distinct developmental lineages that are positioned to receive input from proprioceptor subtypes encoding tibia position, movement, and vibration. 13Bα neurons encode femur-tibia joint angle and mediate postural changes in tibia position. 9Aα neurons also drive changes in leg posture, but encode a combination of directional movement, high frequency vibration, and joint angle. Activating 10Bα neurons, which encode tibia vibration at specific joint angles, elicits pausing in walking flies. Altogether, our results reveal that central circuits integrate information across proprioceptor subtypes to construct complex sensorimotor representations that mediate diverse behaviors, including reflexive control of limb posture and detection of leg vibration.
Keywords: D. melanogaster; behavior; electrophysiology; motor control; neuroscience; proprioception; sensorimotor.
© 2020, Agrawal et al.
Conflict of interest statement
SA, ED, AS, PG, DS, JT, JT No competing interests declared
Figures


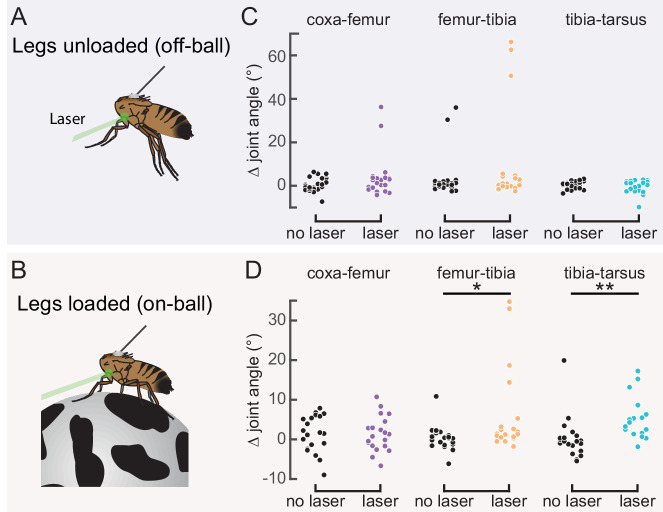
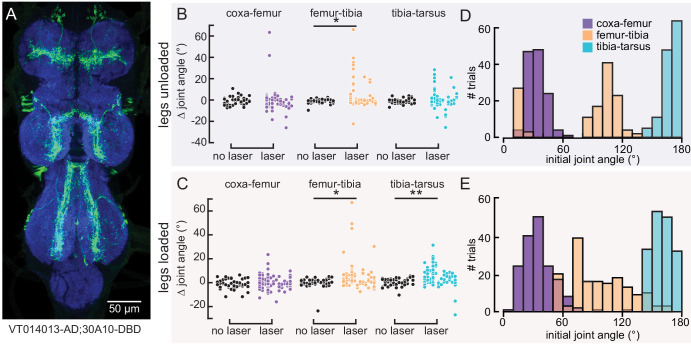
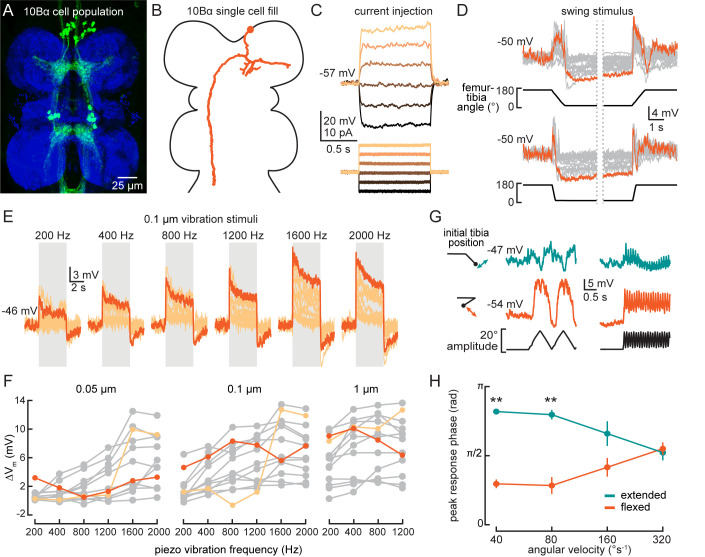
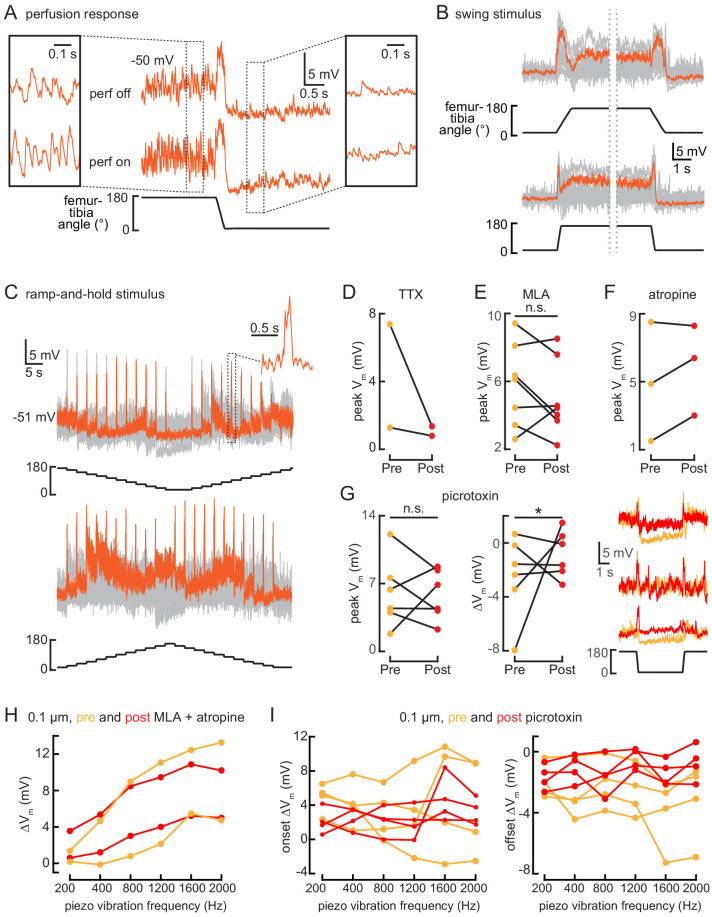
References
Publication types
MeSH terms
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
