

Mapping the electrostatic profiles of cellular membranes
- PMID: 33263429
- PMCID: PMC8098824
- DOI: 10.1091/mbc.E19-08-0436
Mapping the electrostatic profiles of cellular membranes
Abstract
Anionic phospholipids can confer a net negative charge on biological membranes. This surface charge generates an electric field that serves to recruit extrinsic cationic proteins, can alter the disposition of transmembrane proteins and causes the local accumulation of soluble counterions, altering the local pH and the concentration of physiologically important ions such as calcium. Because the phospholipid compositions of the different organellar membranes vary, their surface charges are similarly expected to diverge. Yet, despite the important functional implications, remarkably little is known about the electrostatic properties of the individual organellar membranes. We therefore designed and implemented approaches to estimate the surface charges of the cytosolic membranes of various organelles in situ in intact cells. Our data indicate that the inner leaflet of the plasma membrane is most negative, with a surface potential of approximately -35 mV, followed by the Golgi complex > lysosomes > mitochondria ≈ peroxisomes > endoplasmic reticulum, in decreasing order.
Figures
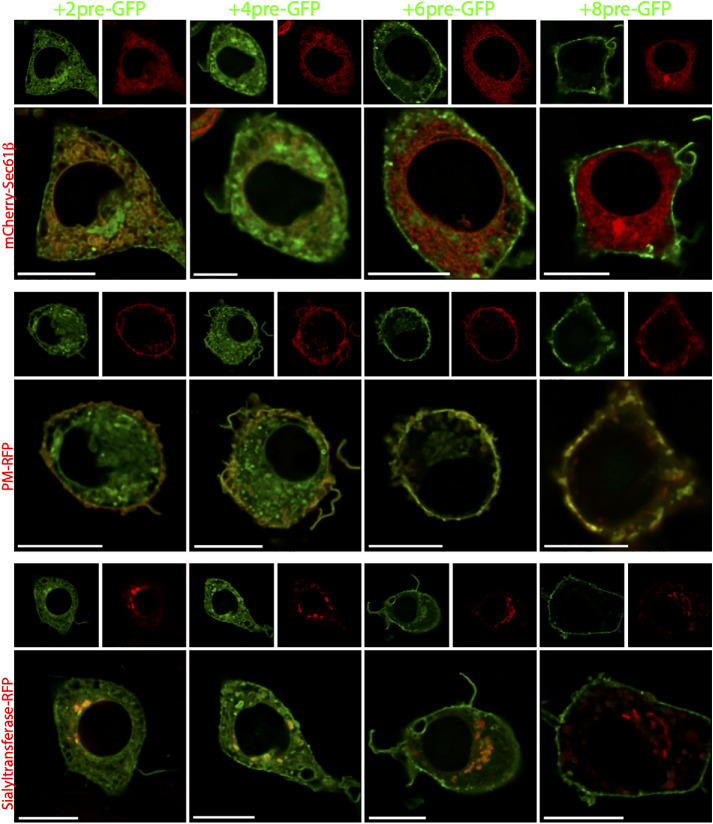




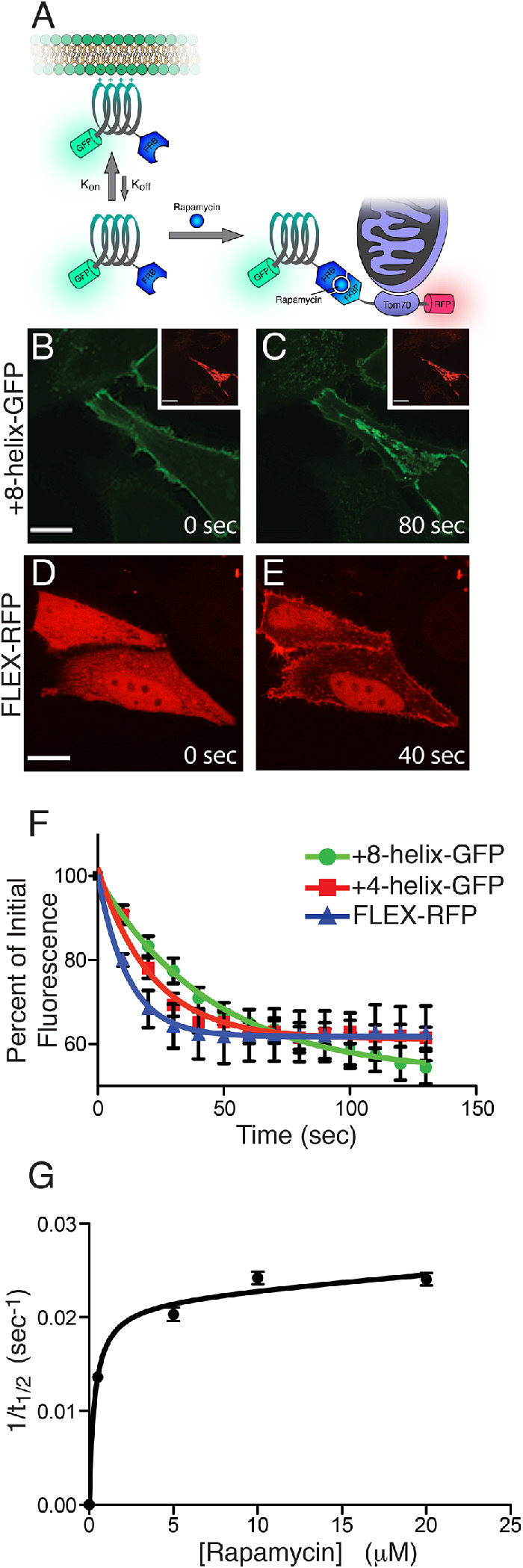
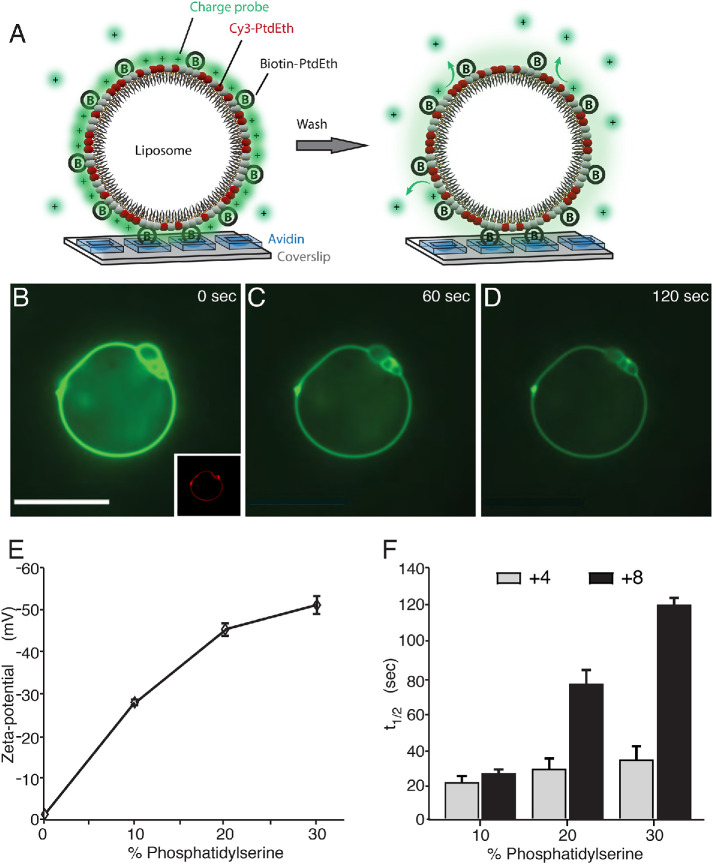
References
-
- Bolte S, Talbot C, Boutte Y, Catrice O, Read ND, Satiat-Jeunemaitre B (2004). FM-dyes as experimental probes for dissecting vesicle trafficking in living plant cells. J Microsc 214, 159–173. - PubMed
-
- Dubyak GR (2007). Go it alone no more—P2X7 joins the society of heteromeric ATP-gated receptor channels. Mol Pharmacol 72, 1402–1405. - PubMed
-
- Ferreira L, Pereira L, Faria R (2015). Fluorescent dyes as a reliable tool in P2X7 receptor-associated pore studies. J Bioenerg Biomembr 47, 283–307. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
