

Optogenetic Tuning Reveals Rho Amplification-Dependent Dynamics of a Cell Contraction Signal Network
- PMID: 33264629
- PMCID: PMC7710677
- DOI: 10.1016/j.celrep.2020.108467
Optogenetic Tuning Reveals Rho Amplification-Dependent Dynamics of a Cell Contraction Signal Network
Abstract
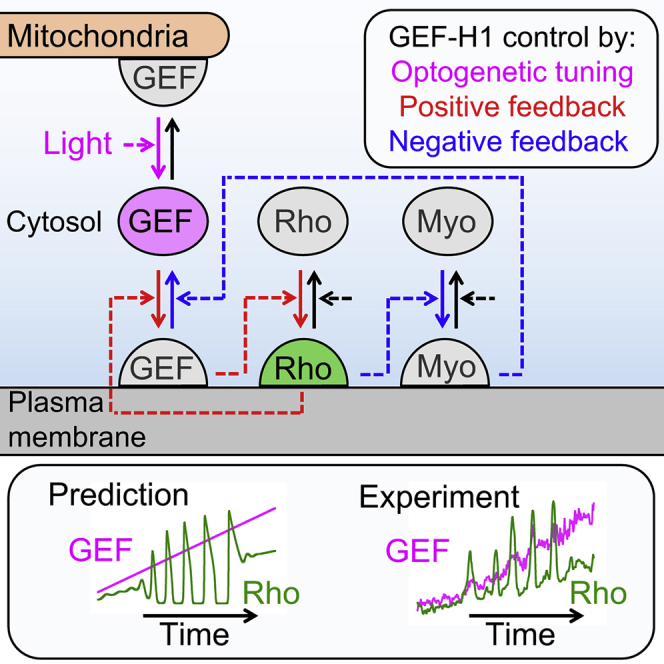
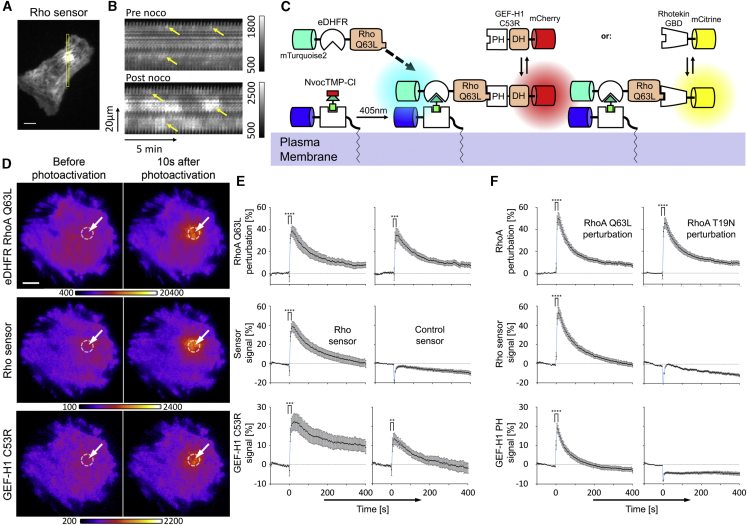
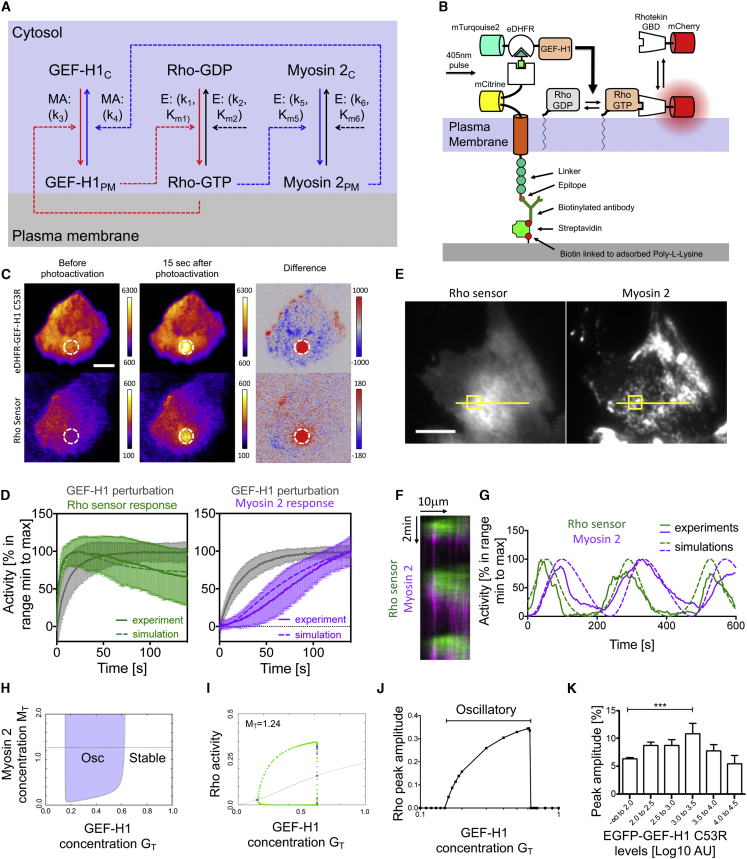
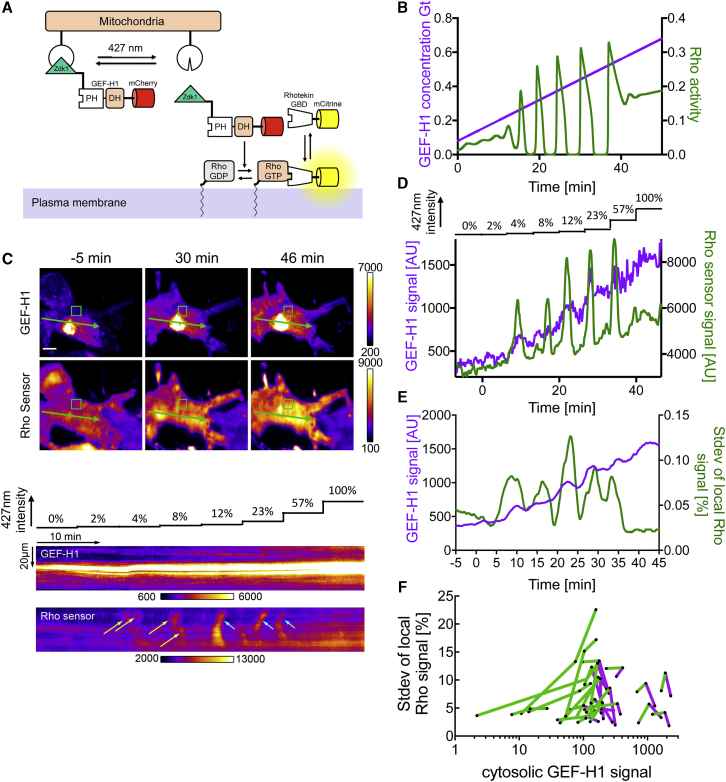
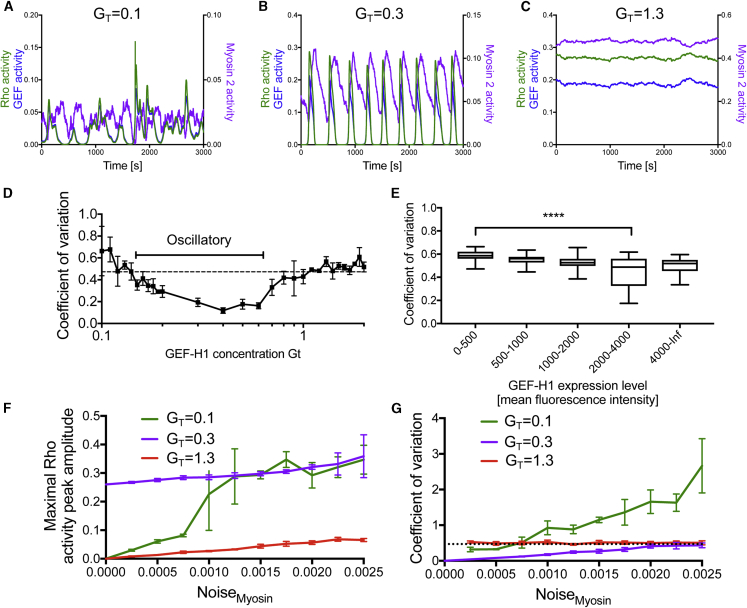
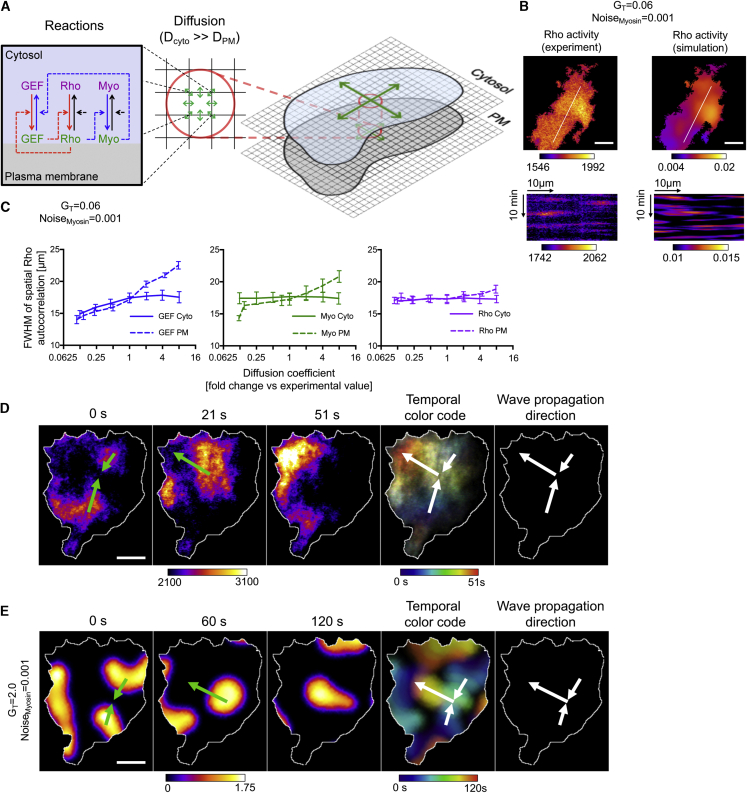
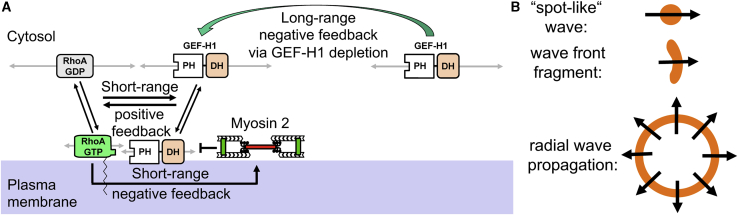
Local cell contraction pulses play important roles in tissue and cell morphogenesis. Here, we improve a chemo-optogenetic approach and apply it to investigate the signal network that generates these pulses. We use these measurements to derive and parameterize a system of ordinary differential equations describing temporal signal network dynamics. Bifurcation analysis and numerical simulations predict a strong dependence of oscillatory system dynamics on the concentration of GEF-H1, an Lbc-type RhoGEF, which mediates the positive feedback amplification of Rho activity. This prediction is confirmed experimentally via optogenetic tuning of the effective GEF-H1 concentration in individual living cells. Numerical simulations show that pulse amplitude is most sensitive to external inputs into the myosin component at low GEF-H1 concentrations and that the spatial pulse width is dependent on GEF-H1 diffusion. Our study offers a theoretical framework to explain the emergence of local cell contraction pulses and their modulation by biochemical and mechanical signals.
Keywords: cell contraction; cytoskeleton; dynamical system; mechanotransduction; myosin; optogenetics; oscillations; parameter inference; reaction-diffusion system; rho GTPase.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
References
-
- Belousov B.P. Периодически действующая реакция и ее механизм [Periodically acting reaction and its mechanism] In: Field R.J., Burger M., editors. Сборник рефератов по радиационной медицине [Collection of Abstracts on Radiation Medicine] Medgiz; 1959. pp. 145–147.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
