

The Role of APOBECs in Viral Replication
- PMID: 33266042
- PMCID: PMC7760323
- DOI: 10.3390/microorganisms8121899
The Role of APOBECs in Viral Replication
Abstract
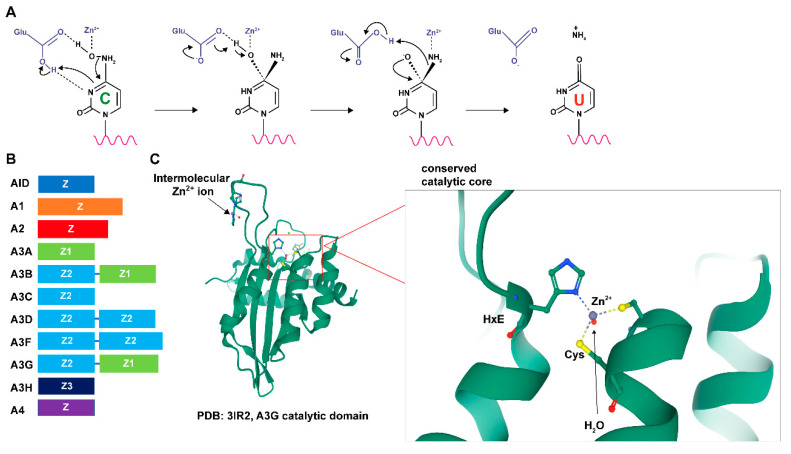
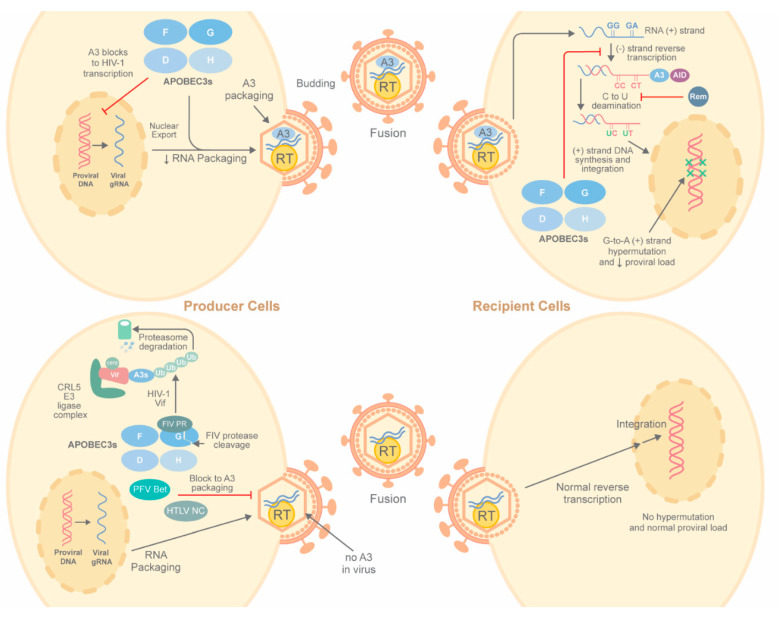
Apolipoprotein B mRNA-editing enzyme catalytic polypeptide-like (APOBEC) proteins are a diverse and evolutionarily conserved family of cytidine deaminases that provide a variety of functions from tissue-specific gene expression and immunoglobulin diversity to control of viruses and retrotransposons. APOBEC family expansion has been documented among mammalian species, suggesting a powerful selection for their activity. Enzymes with a duplicated zinc-binding domain often have catalytically active and inactive domains, yet both have antiviral function. Although APOBEC antiviral function was discovered through hypermutation of HIV-1 genomes lacking an active Vif protein, much evidence indicates that APOBECs also inhibit virus replication through mechanisms other than mutagenesis. Multiple steps of the viral replication cycle may be affected, although nucleic acid replication is a primary target. Packaging of APOBECs into virions was first noted with HIV-1, yet is not a prerequisite for viral inhibition. APOBEC antagonism may occur in viral producer and recipient cells. Signatures of APOBEC activity include G-to-A and C-to-T mutations in a particular sequence context. The importance of APOBEC activity for viral inhibition is reflected in the identification of numerous viral factors, including HIV-1 Vif, which are dedicated to antagonism of these deaminases. Such viral antagonists often are only partially successful, leading to APOBEC selection for viral variants that enhance replication or avoid immune elimination.
Keywords: AID; APOBEC; DNA virus replication; G-to-A mutations; HIV-1 Vif; MMTV Rem; RNA virus replication; genome hypermutation; viral variants.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
