

Functional Identification of Serine Hydroxymethyltransferase as a Key Gene Involved in Lysostaphin Resistance and Virulence Potential of Staphylococcus aureus Strains
- PMID: 33266291
- PMCID: PMC7731198
- DOI: 10.3390/ijms21239135
Functional Identification of Serine Hydroxymethyltransferase as a Key Gene Involved in Lysostaphin Resistance and Virulence Potential of Staphylococcus aureus Strains
Abstract
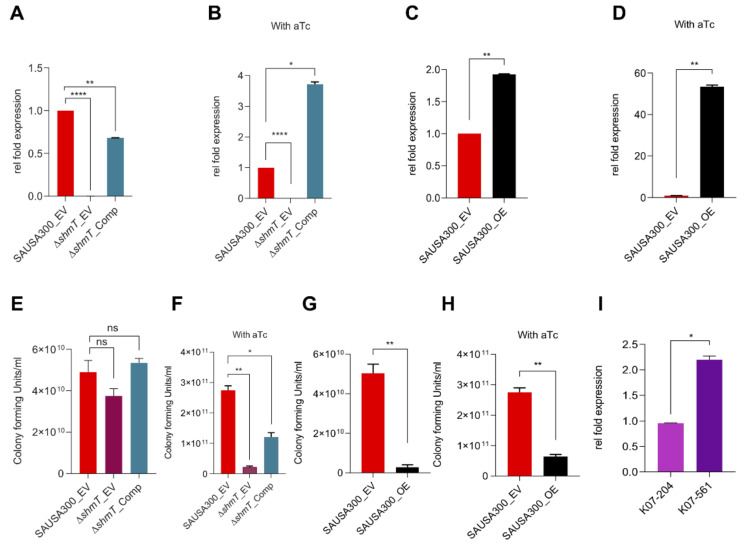
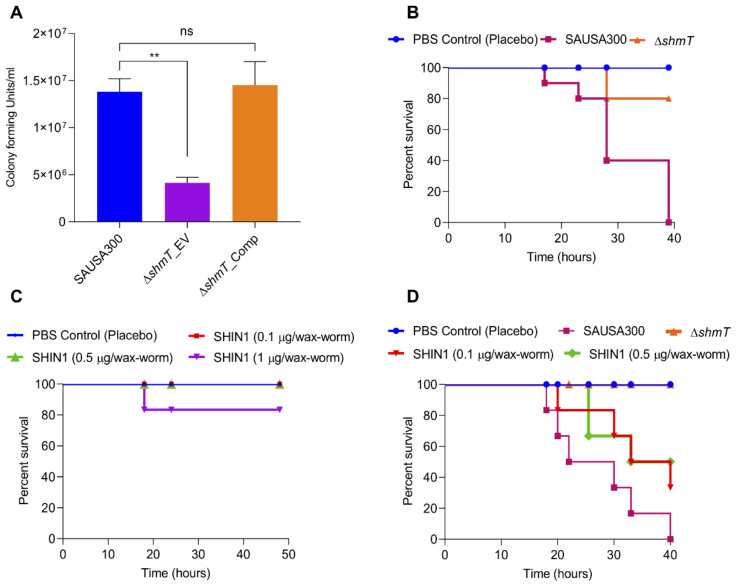
Gaining an insight into the mechanism underlying antimicrobial-resistance development in Staphylococcus aureus is crucial for identifying effective antimicrobials. We isolated S. aureus sequence type 72 from a patient in whom the S. aureus infection was highly resistant to various antibiotics and lysostaphin, but no known resistance mechanisms could explain the mechanism of lysostaphin resistance. Genome-sequencing followed by subtractive and functional genomics revealed that serine hydroxymethyltransferase (glyA or shmT gene) plays a key role in lysostaphin resistance. Serine hydroxymethyltransferase (SHMT) is indispensable for the one-carbon metabolism of serine/glycine interconversion and is linked to folate metabolism. Functional studies revealed the involvement of SHMT in lysostaphin resistance, as ΔshmT was susceptible to the lysostaphin, while complementation of the knockout expressing shmT restored resistance against lysostaphin. In addition, the ΔshmT showed reduced virulence under in vitro (mammalian cell lines infection) and in vivo (wax-worm infection) models. The SHMT inhibitor, serine hydroxymethyltransferase inhibitor 1 (SHIN1), protected the 50% of the wax-worm infected with wild type S. aureus. These results suggest SHMT is relevant to the extreme susceptibility to lysostaphin and the host immune system. Thus, the current study established that SHMT plays a key role in lysostaphin resistance development and in determining the virulence potential of multiple drug-resistant S. aureus.
Keywords: SHMT; SHMT inhibitor; ST72; Staphylococcus aureus; folate cycle; lysostaphin resistance; serine hydroxymethyltransferase; virulence factor.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
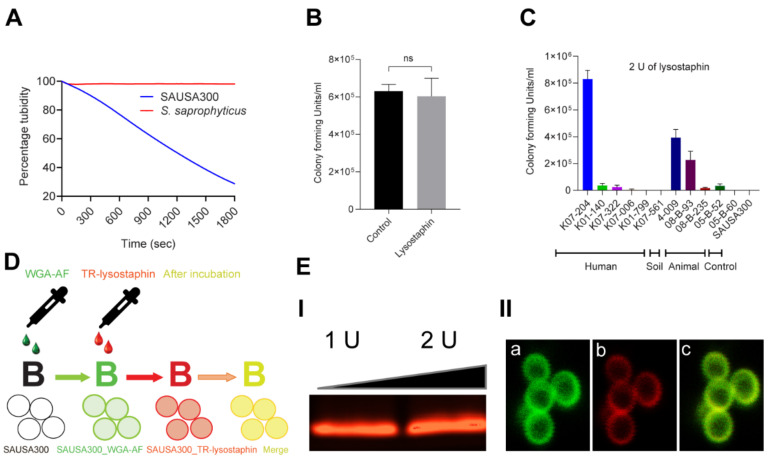
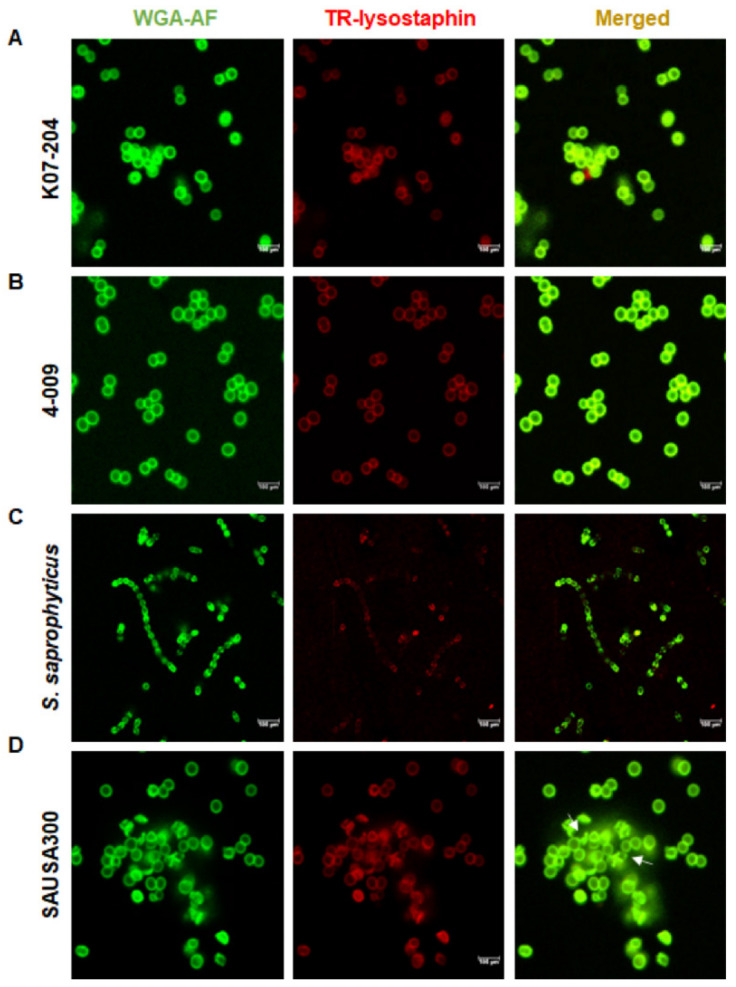
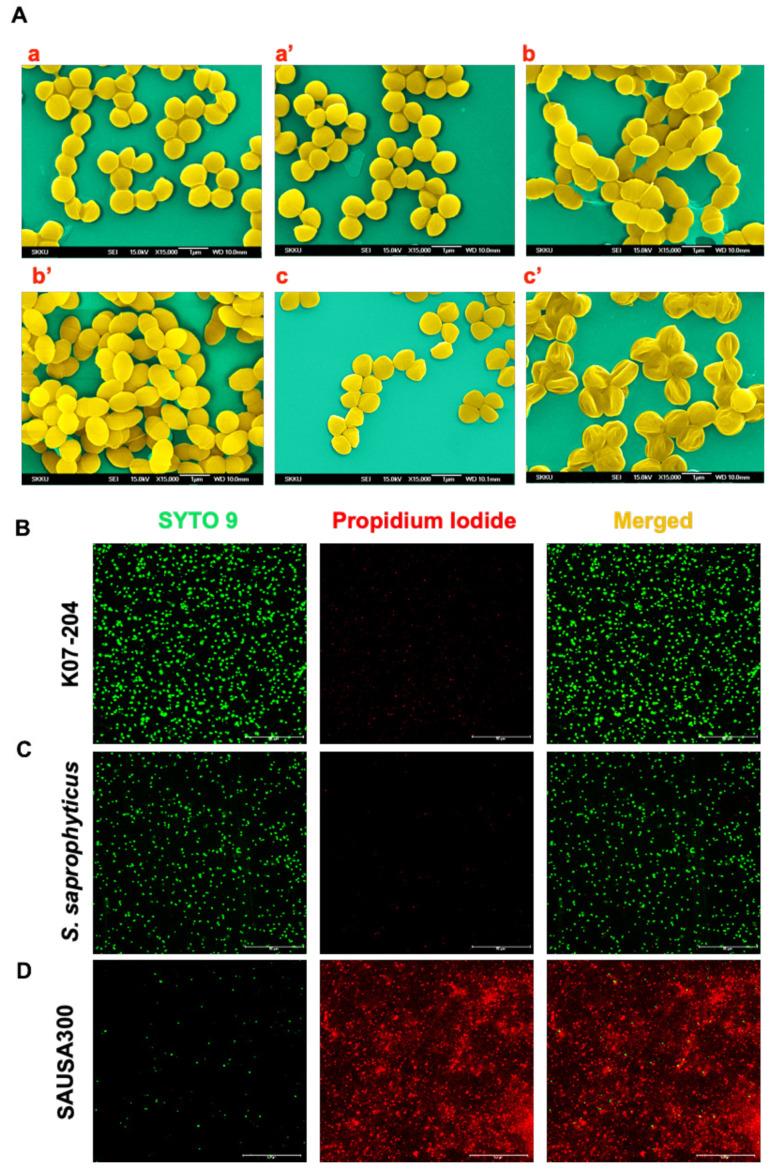
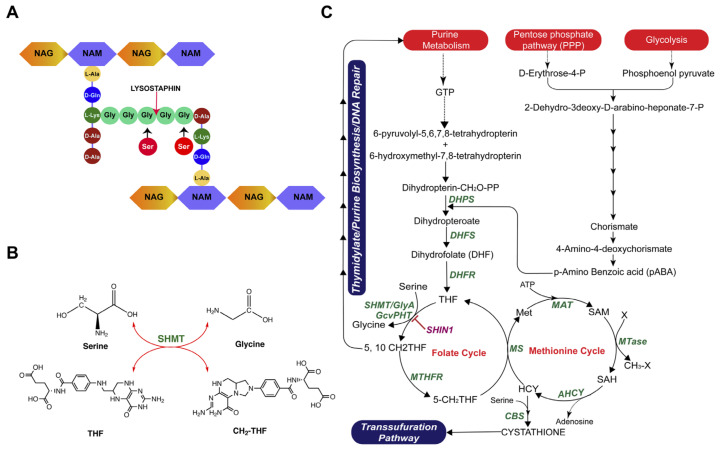
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
