

Priming of SARS-CoV-2 S protein by several membrane-bound serine proteinases could explain enhanced viral infectivity and systemic COVID-19 infection
- PMID: 33268377
- PMCID: PMC7834812
- DOI: 10.1074/jbc.REV120.015980
Priming of SARS-CoV-2 S protein by several membrane-bound serine proteinases could explain enhanced viral infectivity and systemic COVID-19 infection
Abstract
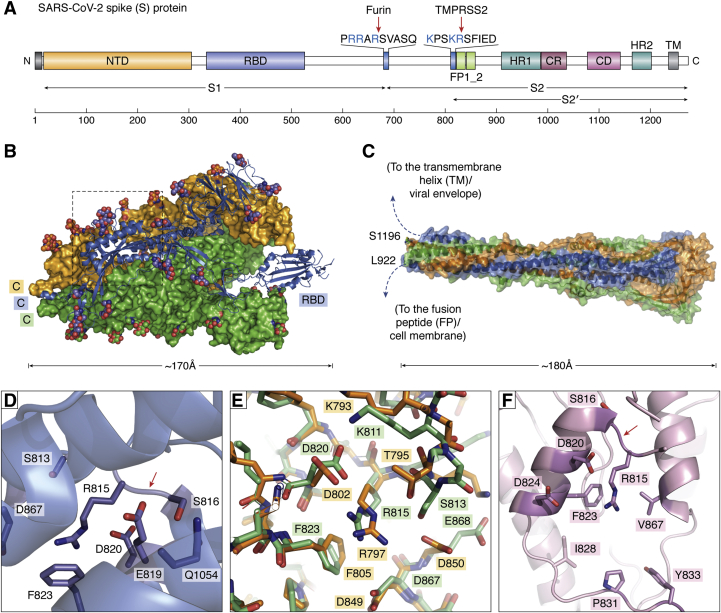
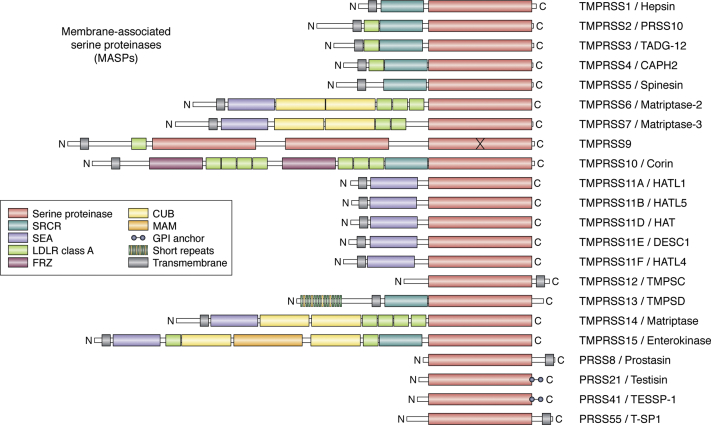
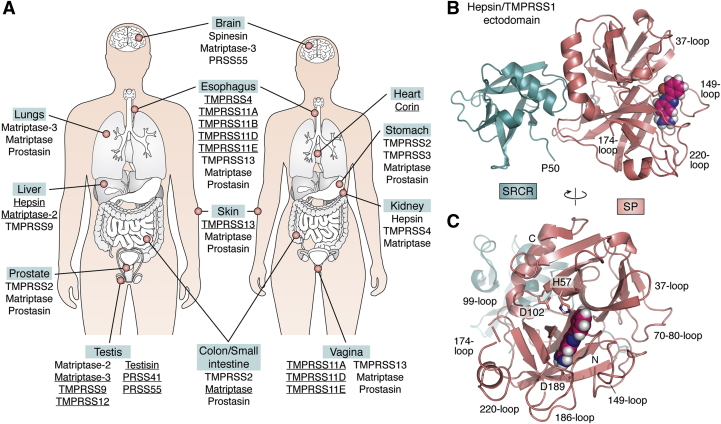
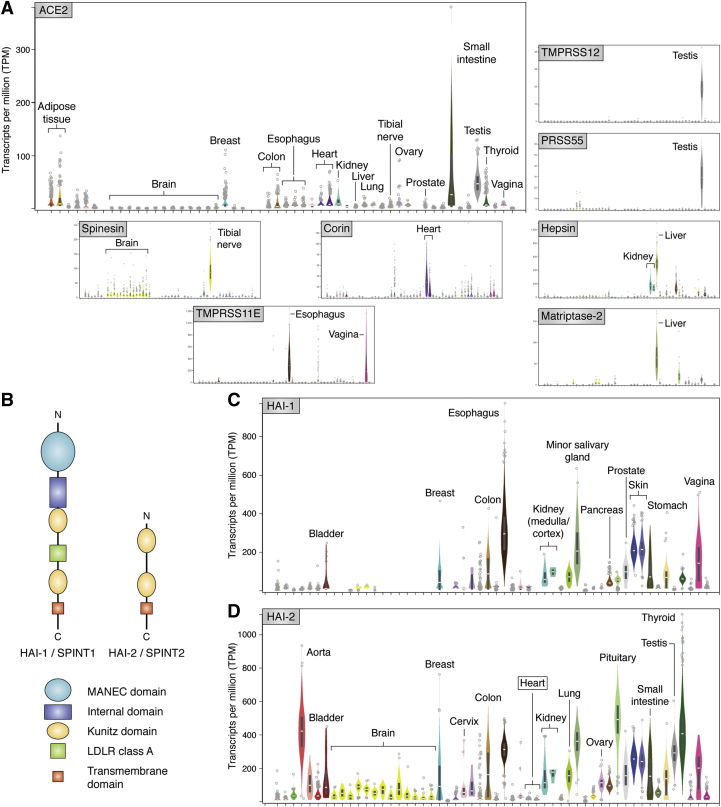
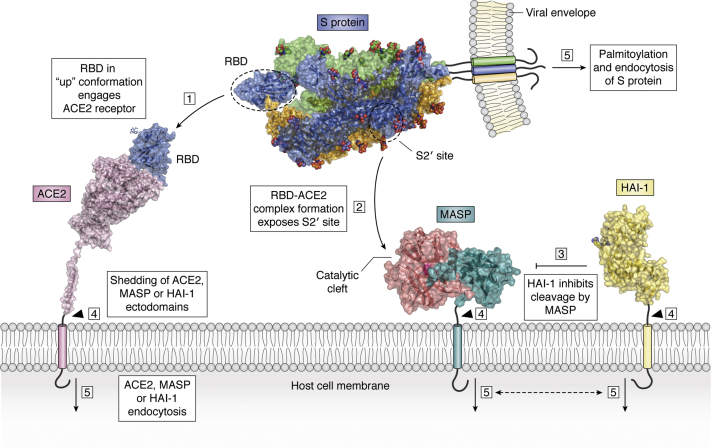
The ongoing COVID-19 pandemic has already caused over a million deaths worldwide, and this death toll will be much higher before effective treatments and vaccines are available. The causative agent of the disease, the coronavirus SARS-CoV-2, shows important similarities with the previously emerged SARS-CoV-1, but also striking differences. First, SARS-CoV-2 possesses a significantly higher transmission rate and infectivity than SARS-CoV-1 and has infected in a few months over 60 million people. Moreover, COVID-19 has a systemic character, as in addition to the lungs, it also affects the heart, liver, and kidneys among other organs of the patients and causes frequent thrombotic and neurological complications. In fact, the term "viral sepsis" has been recently coined to describe the clinical observations. Here I review current structure-function information on the viral spike proteins and the membrane fusion process to provide plausible explanations for these observations. I hypothesize that several membrane-associated serine proteinases (MASPs), in synergy with or in place of TMPRSS2, contribute to activate the SARS-CoV-2 spike protein. Relative concentrations of the attachment receptor, ACE2, MASPs, their endogenous inhibitors (the Kunitz-type transmembrane inhibitors, HAI-1/SPINT1 and HAI-2/SPINT2, as well as major circulating serpins) would determine the infection rate of host cells. The exclusive or predominant expression of major MASPs in specific human organs suggests a direct role of these proteinases in e.g., heart infection and myocardial injury, liver dysfunction, kidney damage, as well as neurological complications. Thorough consideration of these factors could have a positive impact on the control of the current COVID-19 pandemic.
Keywords: COVID-19; HAI-1/SPINT1; TMPRSS2; cell tropism; coronaviruses; membrane-associated serine proteinases (MASPs); serpins; spike (S) protein; structure–function relationship; viral fusion.
Copyright © 2020 The Author. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
Similar articles
-
V367F Mutation in SARS-CoV-2 Spike RBD Emerging during the Early Transmission Phase Enhances Viral Infectivity through Increased Human ACE2 Receptor Binding Affinity.J Virol. 2021 Jul 26;95(16):e0061721. doi: 10.1128/JVI.00617-21. Epub 2021 Jul 26. J Virol. 2021. PMID: 34105996 Free PMC article.
-
Distinctive Roles of Furin and TMPRSS2 in SARS-CoV-2 Infectivity.J Virol. 2022 Apr 27;96(8):e0012822. doi: 10.1128/jvi.00128-22. Epub 2022 Mar 28. J Virol. 2022. PMID: 35343766 Free PMC article.
-
Molecular characterization of interactions between the D614G variant of SARS-CoV-2 S-protein and neutralizing antibodies: A computational approach.Infect Genet Evol. 2021 Jul;91:104815. doi: 10.1016/j.meegid.2021.104815. Epub 2021 Mar 24. Infect Genet Evol. 2021. PMID: 33774178 Free PMC article.
-
Is highly expressed ACE 2 in pregnant women "a curse" in times of COVID-19 pandemic?Life Sci. 2021 Jan 1;264:118676. doi: 10.1016/j.lfs.2020.118676. Epub 2020 Oct 28. Life Sci. 2021. PMID: 33129880 Free PMC article. Review.
-
An Updated Review on Betacoronavirus Viral Entry Inhibitors: Learning from Past Discoveries to Advance COVID-19 Drug Discovery.Curr Top Med Chem. 2021;21(7):571-596. doi: 10.2174/1568026621666210119111409. Curr Top Med Chem. 2021. PMID: 33463470 Review.
Cited by
-
The Physiological TMPRSS2 Inhibitor HAI-2 Alleviates SARS-CoV-2 Infection.J Virol. 2021 May 24;95(12):e00434-21. doi: 10.1128/JVI.00434-21. Print 2021 May 24. J Virol. 2021. PMID: 33789993 Free PMC article. No abstract available.
-
[Impact of SARS-CoV-2/COVID-19 on the placenta].Pathologe. 2021 Nov;42(6):591-597. doi: 10.1007/s00292-021-00952-7. Epub 2021 Jun 11. Pathologe. 2021. PMID: 34115179 Free PMC article. Review. German.
-
Enoxaparin augments alpha-1-antitrypsin inhibition of TMPRSS2, a promising drug combination against COVID-19.Sci Rep. 2022 Mar 25;12(1):5207. doi: 10.1038/s41598-022-09133-9. Sci Rep. 2022. PMID: 35338216 Free PMC article.
-
Millisecond dynamic of SARS-CoV-2 spike and its interaction with ACE2 receptor and small extracellular vesicles.J Extracell Vesicles. 2021 Dec;10(14):e12170. doi: 10.1002/jev2.12170. J Extracell Vesicles. 2021. PMID: 34874124 Free PMC article.
-
Natural variants in SARS-CoV-2 Spike protein pinpoint structural and functional hotspots with implications for prophylaxis and therapeutic strategies.Sci Rep. 2021 Jun 23;11(1):13120. doi: 10.1038/s41598-021-92641-x. Sci Rep. 2021. PMID: 34162970 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
