

Glycine-based treatment ameliorates NAFLD by modulating fatty acid oxidation, glutathione synthesis, and the gut microbiome
- PMID: 33268508
- PMCID: PMC7982985
- DOI: 10.1126/scitranslmed.aaz2841
Glycine-based treatment ameliorates NAFLD by modulating fatty acid oxidation, glutathione synthesis, and the gut microbiome
Abstract
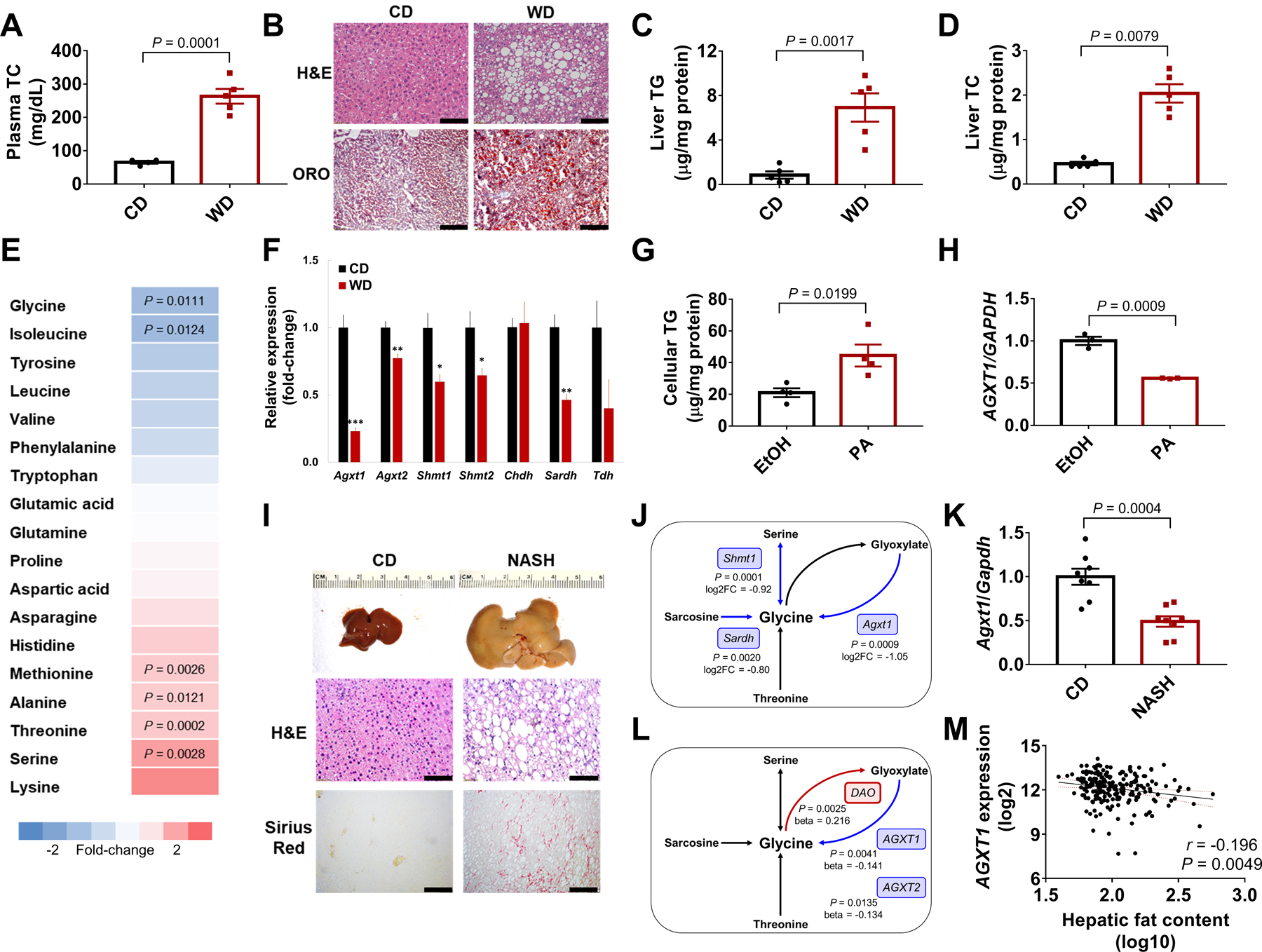
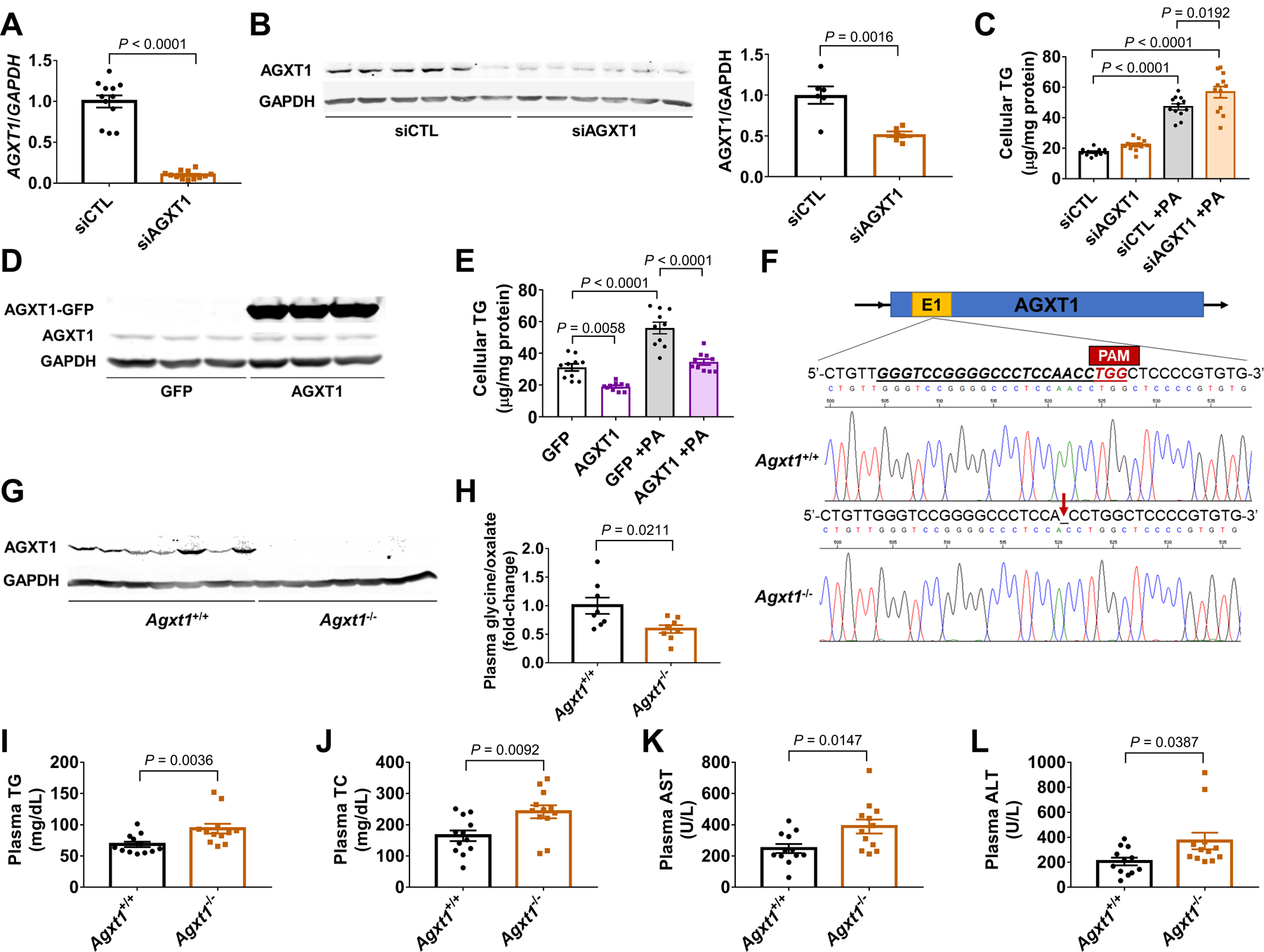
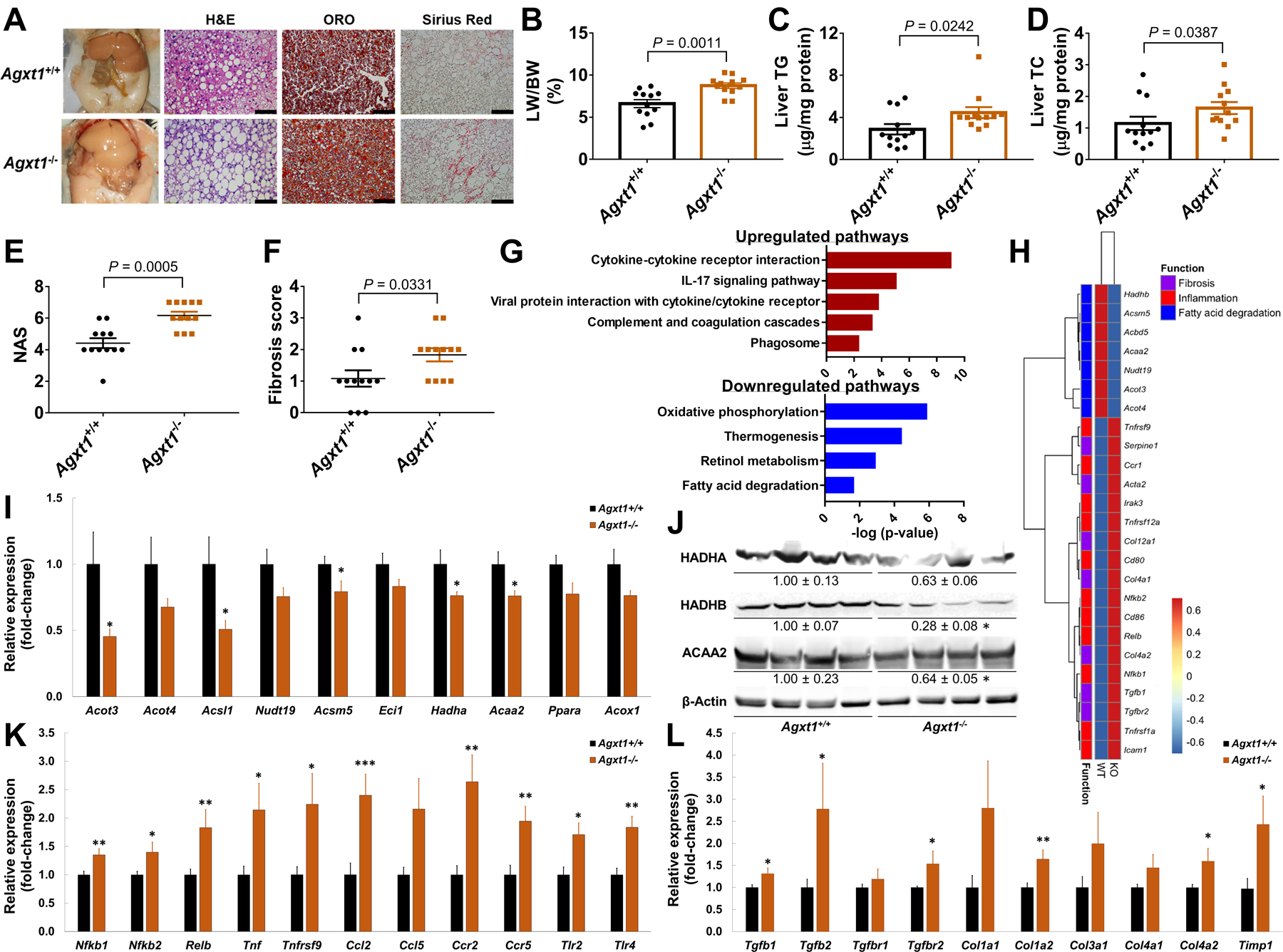
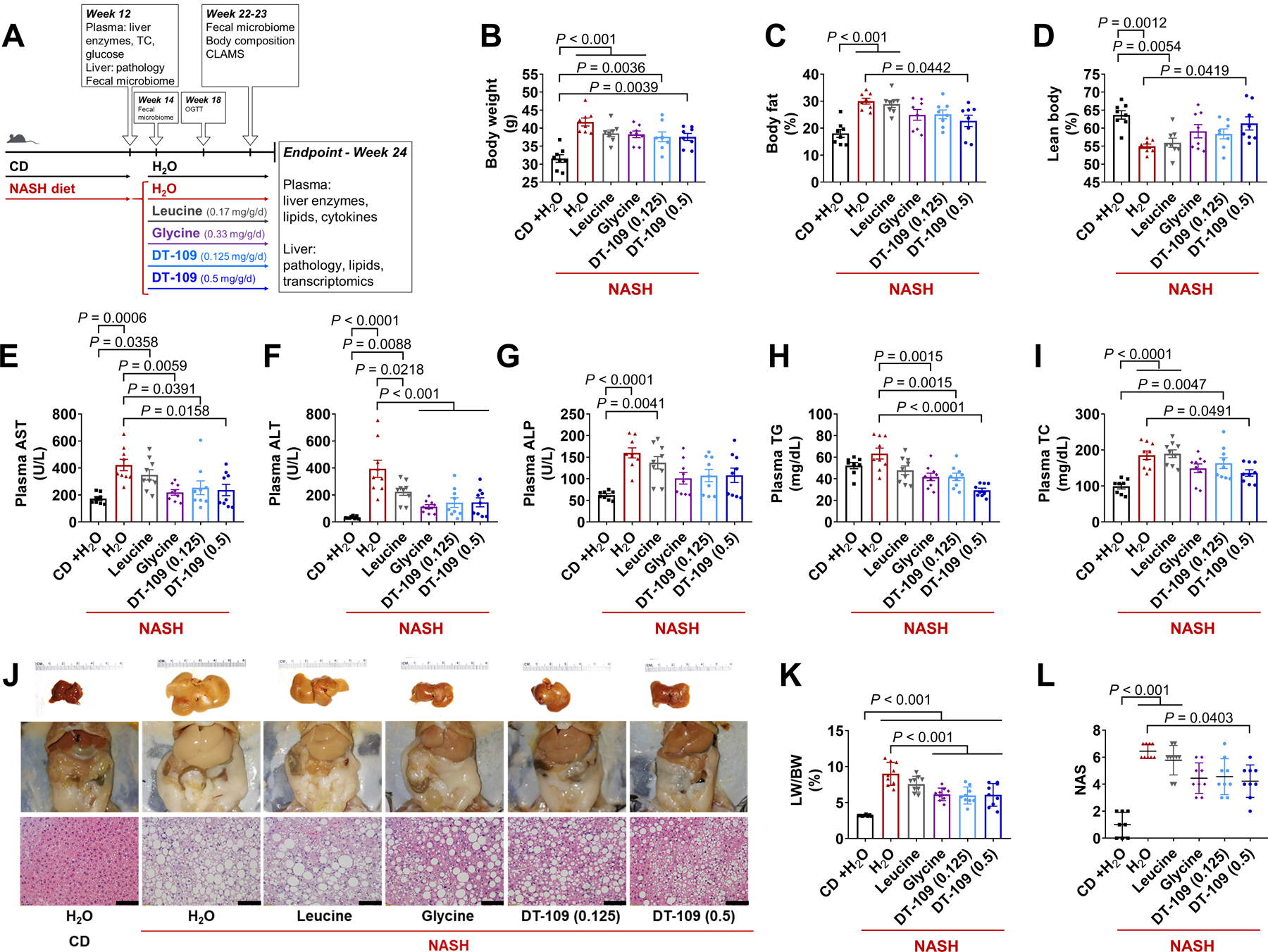
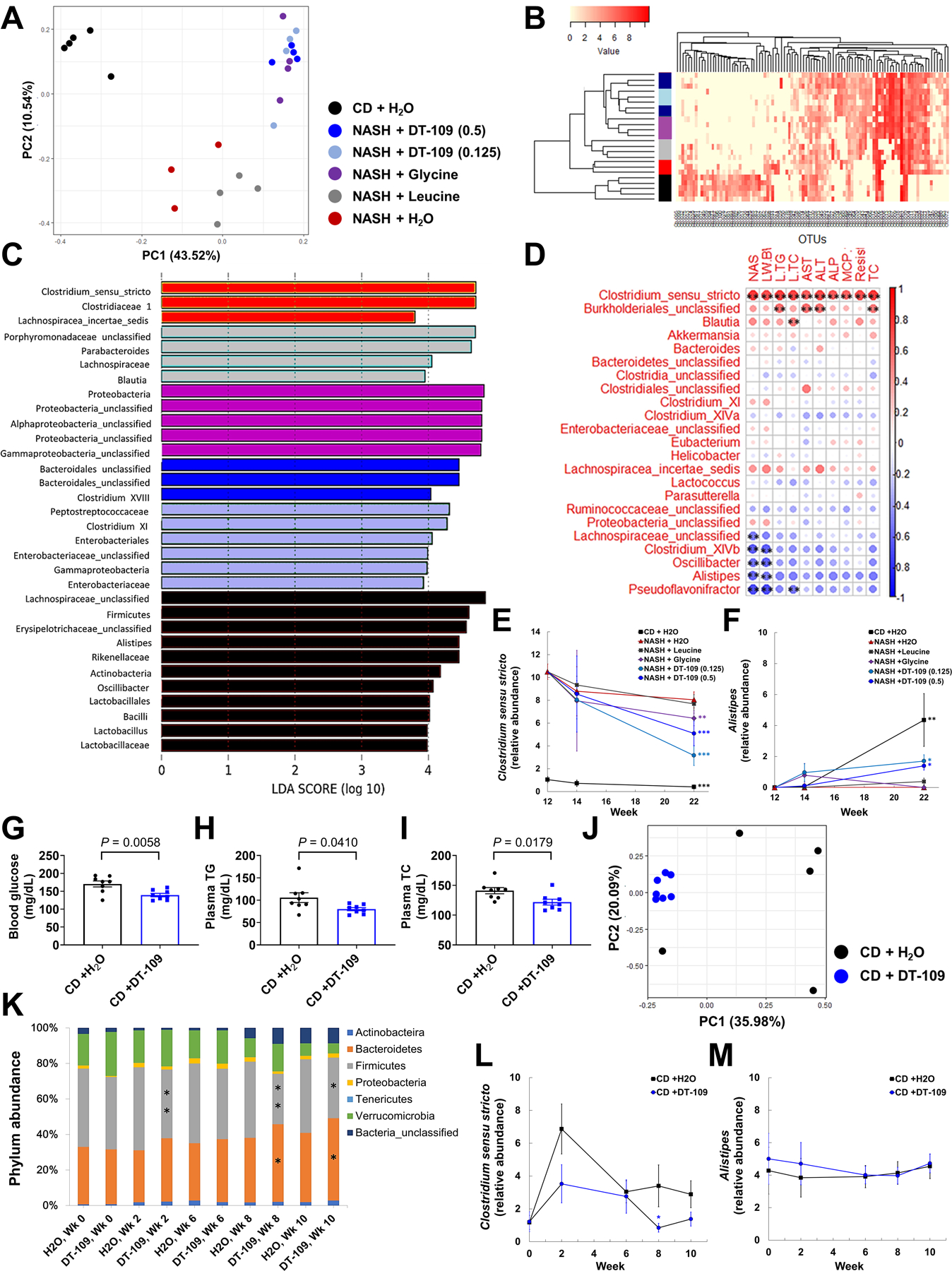
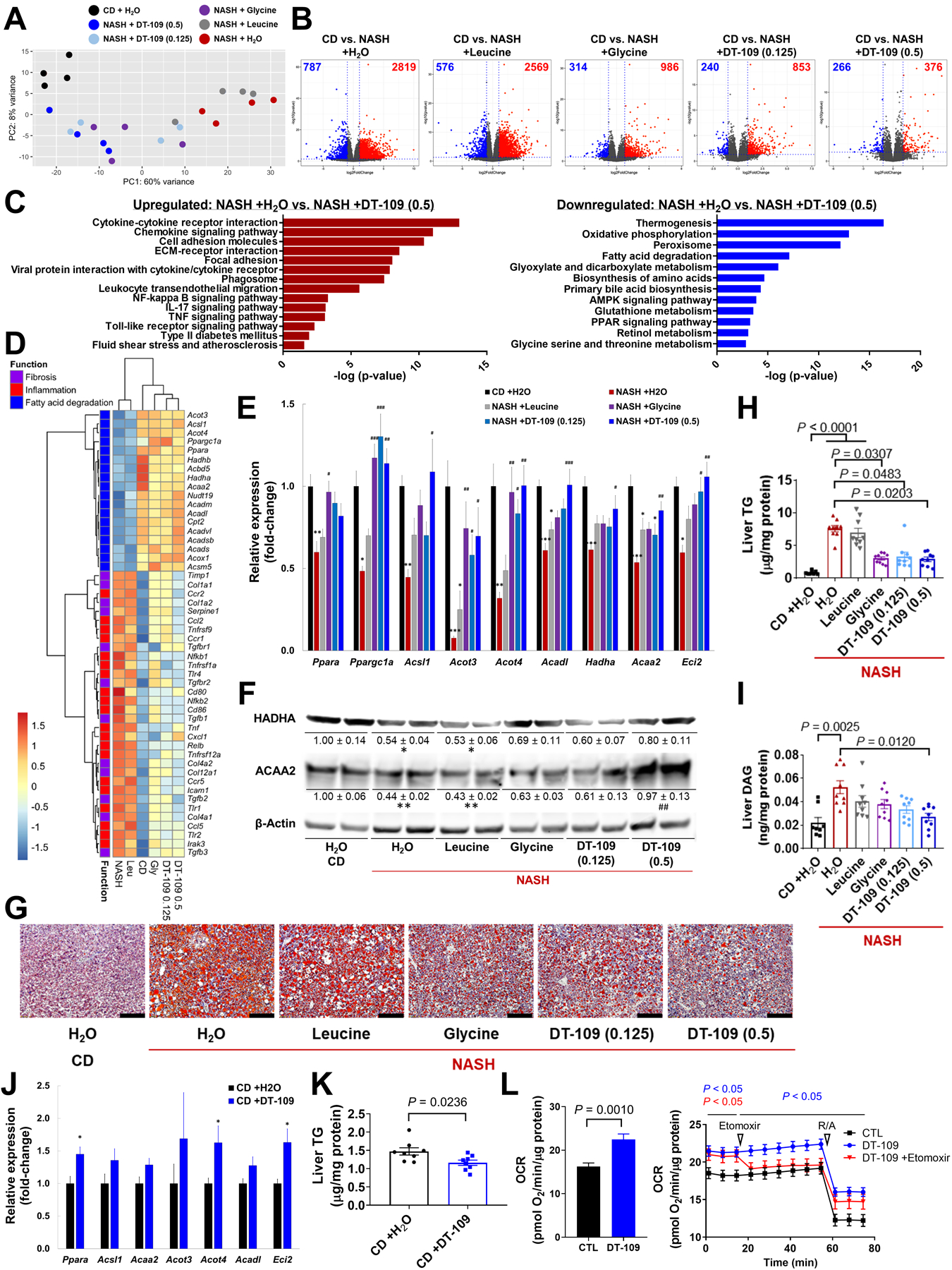
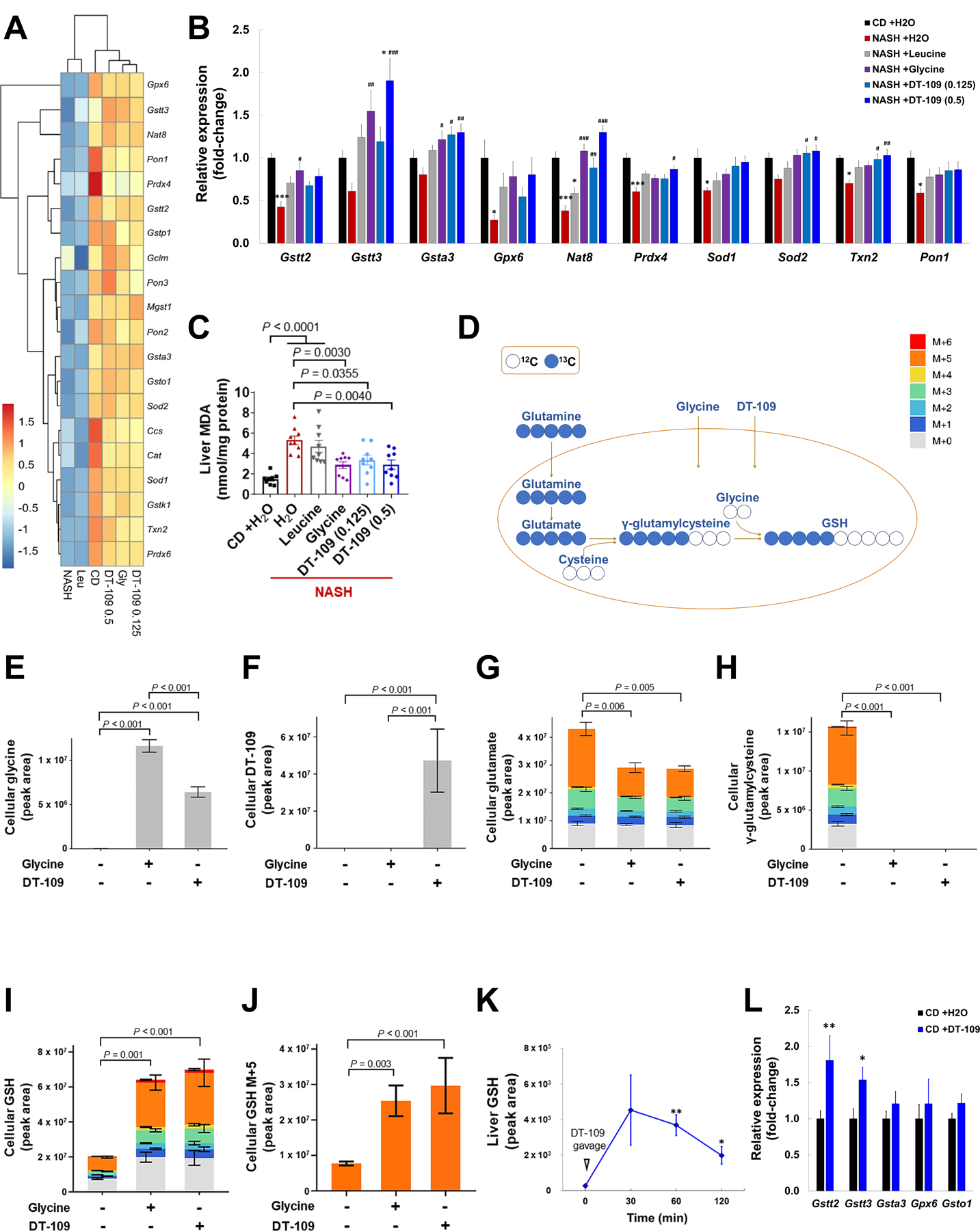
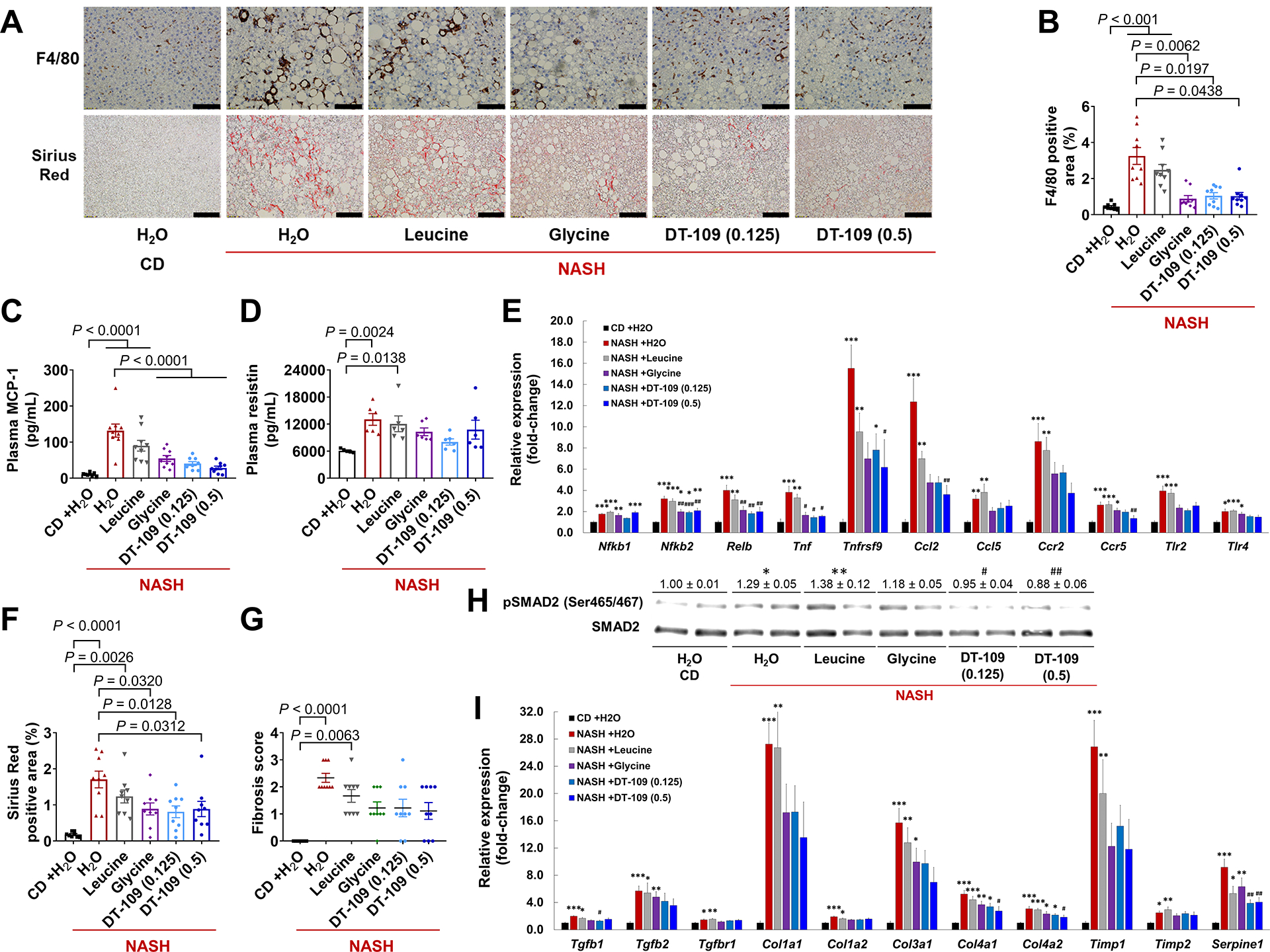
Nonalcoholic fatty liver disease (NAFLD) including nonalcoholic steatohepatitis (NASH) has reached epidemic proportions with no pharmacological therapy approved. Lower circulating glycine is consistently reported in patients with NAFLD, but the causes for reduced glycine, its role as a causative factor, and its therapeutic potential remain unclear. We performed transcriptomics in livers from humans and mice with NAFLD and found suppression of glycine biosynthetic genes, primarily alanine-glyoxylate aminotransferase 1 (AGXT1). Genetic (Agxt1 -/- mice) and dietary approaches to limit glycine availability resulted in exacerbated diet-induced hyperlipidemia and steatohepatitis, with suppressed mitochondrial/peroxisomal fatty acid β-oxidation (FAO) and enhanced inflammation as the underlying pathways. We explored glycine-based compounds with dual lipid/glucose-lowering properties as potential therapies for NAFLD and identified a tripeptide (Gly-Gly-L-Leu, DT-109) that improved body composition and lowered circulating glucose, lipids, transaminases, proinflammatory cytokines, and steatohepatitis in mice with established NASH induced by a high-fat, cholesterol, and fructose diet. We applied metagenomics, transcriptomics, and metabolomics to explore the underlying mechanisms. The bacterial genus Clostridium sensu stricto was markedly increased in mice with NASH and decreased after DT-109 treatment. DT-109 induced hepatic FAO pathways, lowered lipotoxicity, and stimulated de novo glutathione synthesis. In turn, inflammatory infiltration and hepatic fibrosis were attenuated via suppression of NF-κB target genes and TGFβ/SMAD signaling. Unlike its effects on the gut microbiome, DT-109 stimulated FAO and glutathione synthesis independent of NASH. In conclusion, impaired glycine metabolism may play a causative role in NAFLD. Glycine-based treatment attenuates experimental NAFLD by stimulating hepatic FAO and glutathione synthesis, thus warranting clinical evaluation.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
References
-
- Younossi ZM, Koenig AB, Abdelatif D, Fazel Y, Henry L, Wymer M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology. 64, 73–84 (2016). - PubMed
-
- Bedossa P, Poitou C, Veyrie N, Bouillot JL, Basdevant A, Paradis V, Tordjman J, Clement K. Histopathological algorithm and scoring system for evaluation of liver lesions in morbidly obese patients. Hepatology. 56, 1751–17599 (2012). - PubMed
-
- Sinn DH, Cho SJ, Gu S, Seong D, Kang D, Kim H, Yi BK, Paik SW, Guallar E, Cho J, Gwak GY. Persistent Nonalcoholic Fatty Liver Disease Increases Risk for Carotid Atherosclerosis. Gastroenterology. 151, 481–488.e1 (2016). - PubMed
-
- Younes R, Bugianesi E. A spotlight on pathogenesis, interactions and novel therapeutic options in NAFLD. Nat. Rev. Gastroenterol. Hepatol 16,80–82 (2019). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- K99 HL150233/HL/NHLBI NIH HHS/United States
- R01 HL123333/HL/NHLBI NIH HHS/United States
- P30 DK089503/DK/NIDDK NIH HHS/United States
- P60 DK020572/DK/NIDDK NIH HHS/United States
- R01 HL134569/HL/NHLBI NIH HHS/United States
- P30 DK020572/DK/NIDDK NIH HHS/United States
- U2C DK110768/DK/NIDDK NIH HHS/United States
- R00 HL150233/HL/NHLBI NIH HHS/United States
- R01 DK106540/DK/NIDDK NIH HHS/United States
- R01 HL138139/HL/NHLBI NIH HHS/United States
- R01 HL137214/HL/NHLBI NIH HHS/United States
- P30 CA046592/CA/NCI NIH HHS/United States
- R01 HL068878/HL/NHLBI NIH HHS/United States
- R01 HL145176/HL/NHLBI NIH HHS/United States
- U24 DK097153/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
