

Artificial consortium demonstrates emergent properties of enhanced cellulosic-sugar degradation and biofuel synthesis
- PMID: 33268782
- PMCID: PMC7710750
- DOI: 10.1038/s41522-020-00170-8
Artificial consortium demonstrates emergent properties of enhanced cellulosic-sugar degradation and biofuel synthesis
Abstract
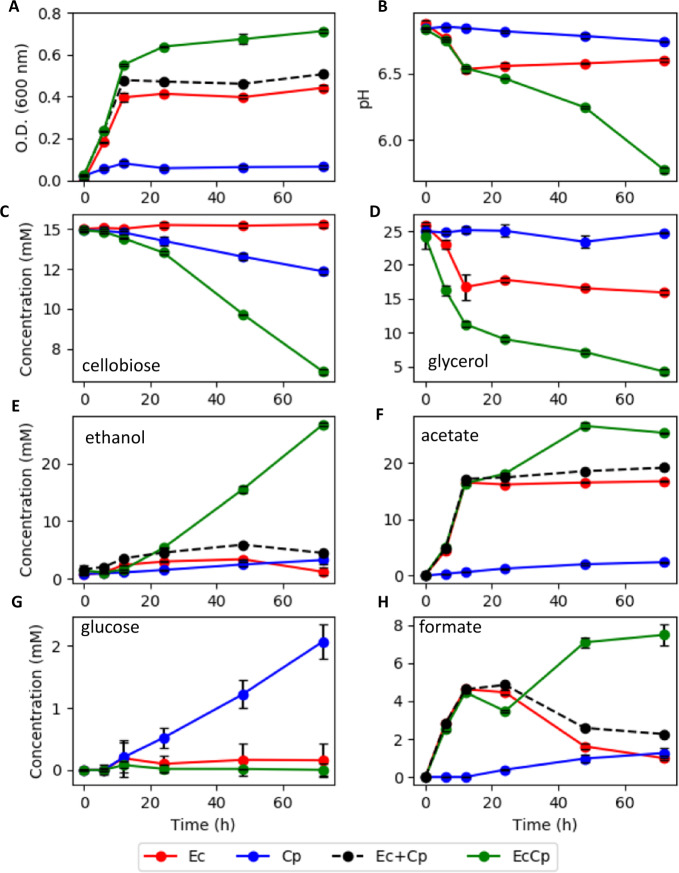
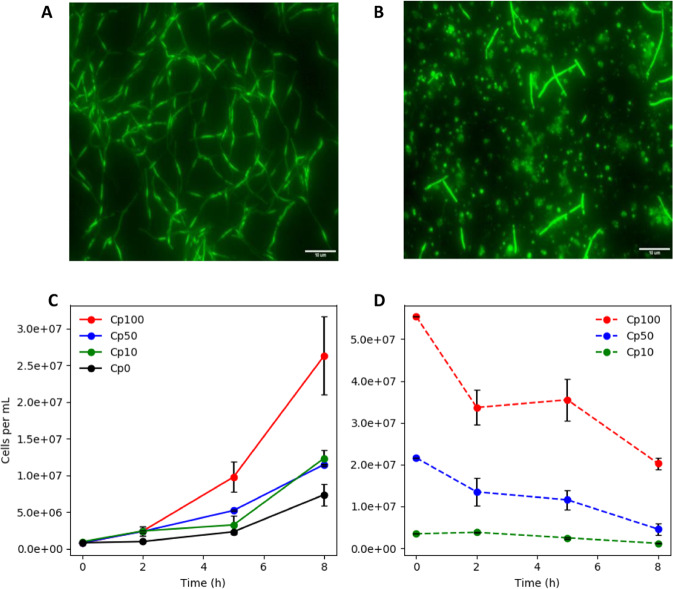
Planktonic cultures, of a rationally designed consortium, demonstrated emergent properties that exceeded the sums of monoculture properties, including a >200% increase in cellobiose catabolism, a >100% increase in glycerol catabolism, a >800% increase in ethanol production, and a >120% increase in biomass productivity. The consortium was designed to have a primary and secondary-resource specialist that used crossfeeding with a positive feedback mechanism, division of labor, and nutrient and energy transfer via necromass catabolism. The primary resource specialist was Clostridium phytofermentans (a.k.a. Lachnoclostridium phytofermentans), a cellulolytic, obligate anaerobe. The secondary-resource specialist was Escherichia coli, a versatile, facultative anaerobe, which can ferment glycerol and byproducts of cellobiose catabolism. The consortium also demonstrated emergent properties of enhanced biomass accumulation when grown as biofilms, which created high cell density communities with gradients of species along the vertical axis. Consortium biofilms were robust to oxic perturbations with E. coli consuming O2, creating an anoxic environment for C. phytofermentans. Anoxic/oxic cycling further enhanced biomass productivity of the biofilm consortium, increasing biomass accumulation ~250% over the sum of the monoculture biofilms. Consortium emergent properties were credited to several synergistic mechanisms. E. coli consumed inhibitory byproducts from cellobiose catabolism, driving higher C. phytofermentans growth and higher cellulolytic enzyme production, which in turn provided more substrate for E. coli. E. coli necromass enhanced C. phytofermentans growth while C. phytofermentans necromass aided E. coli growth via the release of peptides and amino acids, respectively. In aggregate, temporal cycling of necromass constituents increased flux of cellulose-derived resources through the consortium. The study establishes a consortia-based, bioprocessing strategy built on naturally occurring interactions for improved conversion of cellulose-derived sugars into bioproducts.
Conflict of interest statement
The authors declare no competing interests.
Figures







References
-
- Yang B, Wyman CE. Pretreatment: the key to unlocking low-cost cellulosic ethanol. Biofuels Bioprod. Bioref. 2008;2:26–40. doi: 10.1002/bbb.49. - DOI
-
- Mohite, B. V. & Patil, S. V. in New and Future Developments in Microbial Biotechnology and Bioengineering (ed. Gupta, V. K.) Ch. 4, 31–40 (Elsevier, 2016).
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
