

This is a preprint.
A monoclonal antibody against staphylococcal enterotoxin B superantigen inhibits SARS-CoV-2 entry in vitro
- PMID: 33269352
- PMCID: PMC7709177
- DOI: 10.1101/2020.11.24.395079
A monoclonal antibody against staphylococcal enterotoxin B superantigen inhibits SARS-CoV-2 entry in vitro
Update in
-
A monoclonal antibody against staphylococcal enterotoxin B superantigen inhibits SARS-CoV-2 entry in vitro.Structure. 2021 Sep 2;29(9):951-962.e3. doi: 10.1016/j.str.2021.04.005. Epub 2021 Apr 29. Structure. 2021. PMID: 33930306 Free PMC article.
Abstract
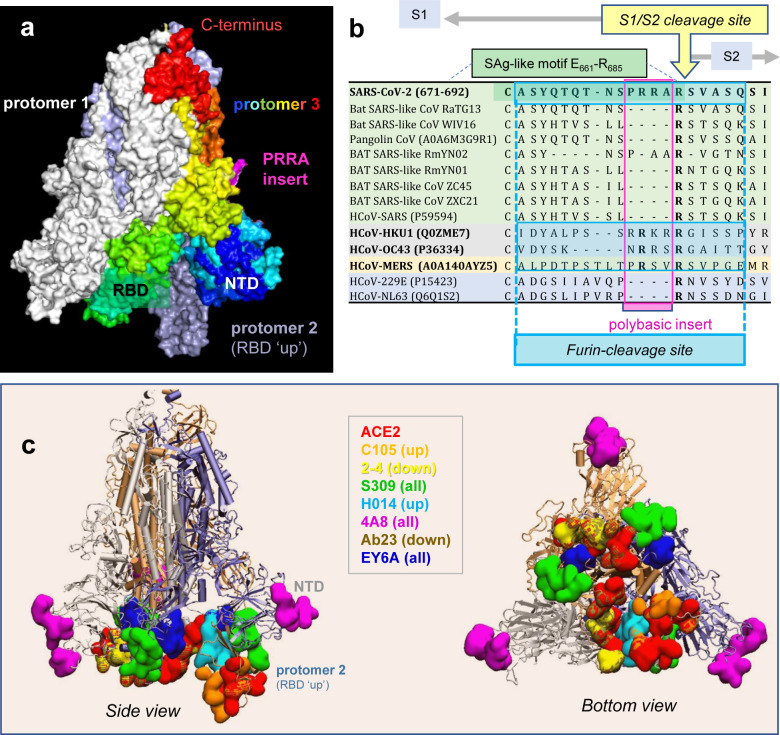
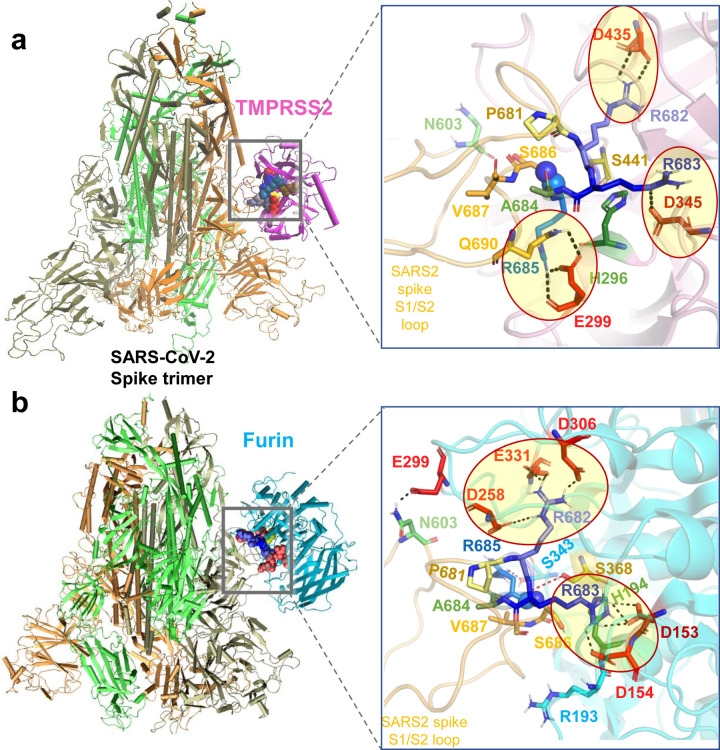
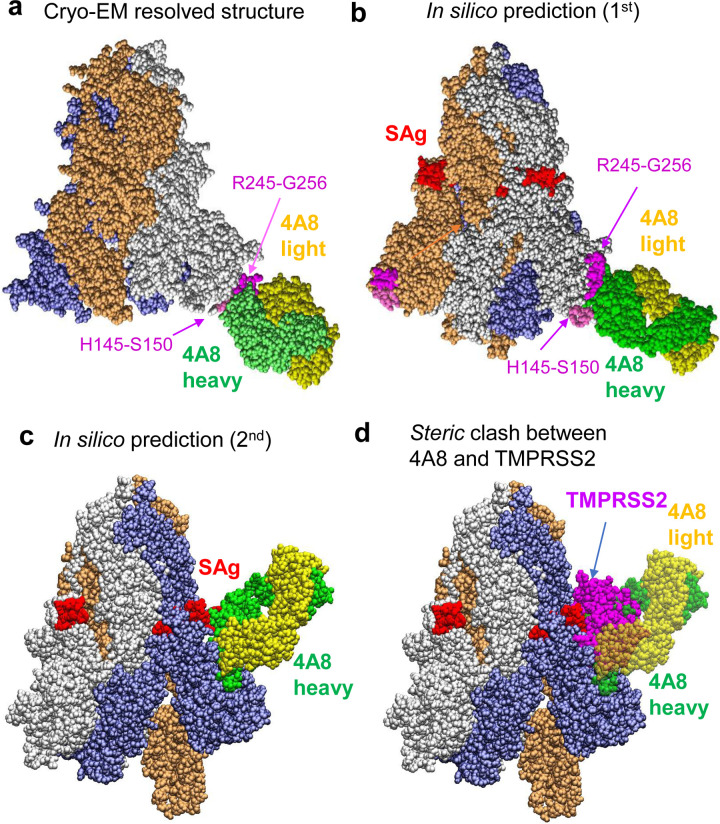
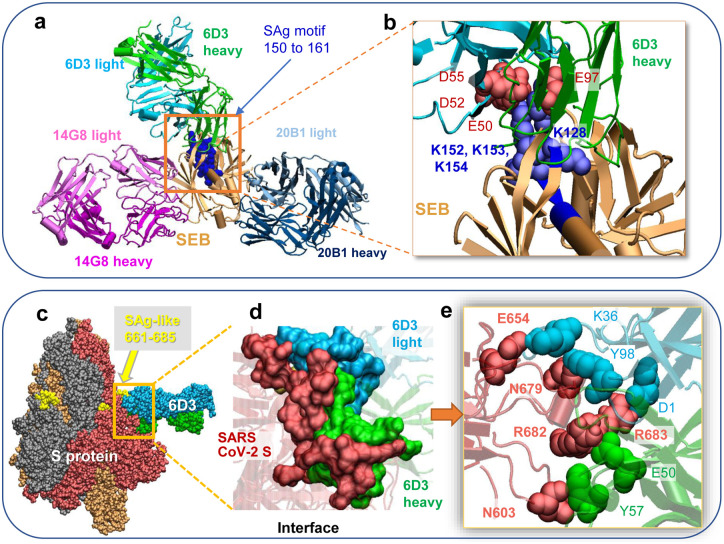
We recently discovered a superantigen-like motif, similar to Staphylococcal enterotoxin B (SEB), near the S1/S2 cleavage site of SARS-CoV-2 Spike protein, which might explain the multisystem-inflammatory syndrome (MIS-C) observed in children and cytokine storm in severe COVID-19 patients. We show here that an anti-SEB monoclonal antibody (mAb), 6D3, can bind this viral motif, and in particular its PRRA insert, to inhibit infection by blocking the access of host cell proteases, TMPRSS2 or furin, to the cleavage site. The high affinity of 6D3 for the furin-cleavage site originates from a poly-acidic segment at its heavy chain CDR2, a feature shared with SARS-CoV-2-neutralizing mAb 4A8. The affinity of 6D3 and 4A8 for this site points to their potential utility as therapeutics for treating COVID-19, MIS-C, or common cold caused by human coronaviruses (HCoVs) that possess a furin-like cleavage site.
Conflict of interest statement
Declarations of Interest
The authors have nothing to declare.
Figures

References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous