

Interaction of transcription factor AP-2 gamma with proto-oncogene PELP1 promotes tumorigenesis by enhancing RET signaling
- PMID: 33269540
- PMCID: PMC8024722
- DOI: 10.1002/1878-0261.12871
Interaction of transcription factor AP-2 gamma with proto-oncogene PELP1 promotes tumorigenesis by enhancing RET signaling
Erratum in
-
Corrigendum to: Interaction of transcription factor AP-2 gamma with proto-oncogene PELP1 promotes tumorigenesis by enhancing RET signaling.Mol Oncol. 2022 Jan;16(1):307. doi: 10.1002/1878-0261.13129. Epub 2021 Nov 10. Mol Oncol. 2022. PMID: 34755450 Free PMC article. No abstract available.
Abstract
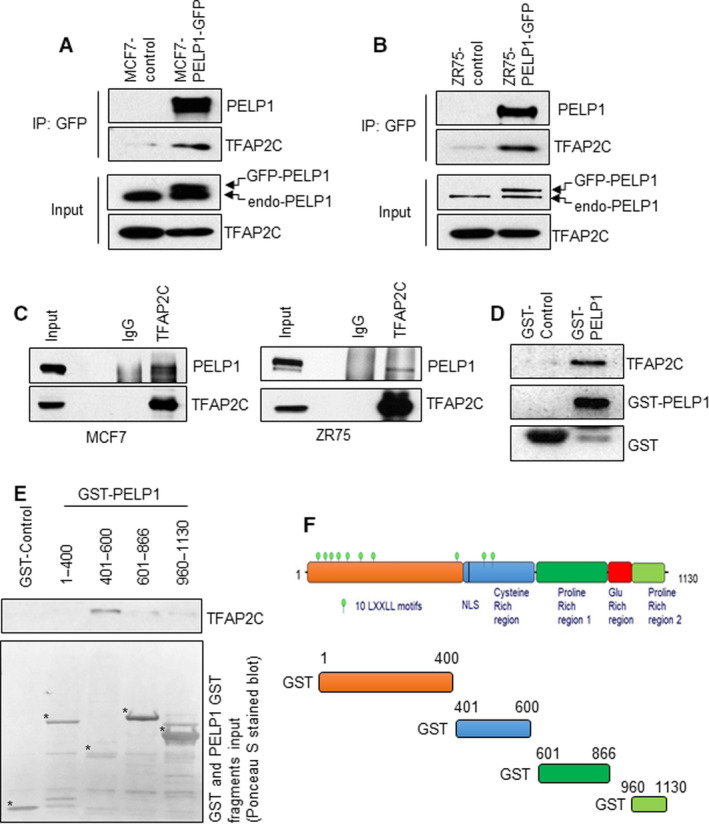
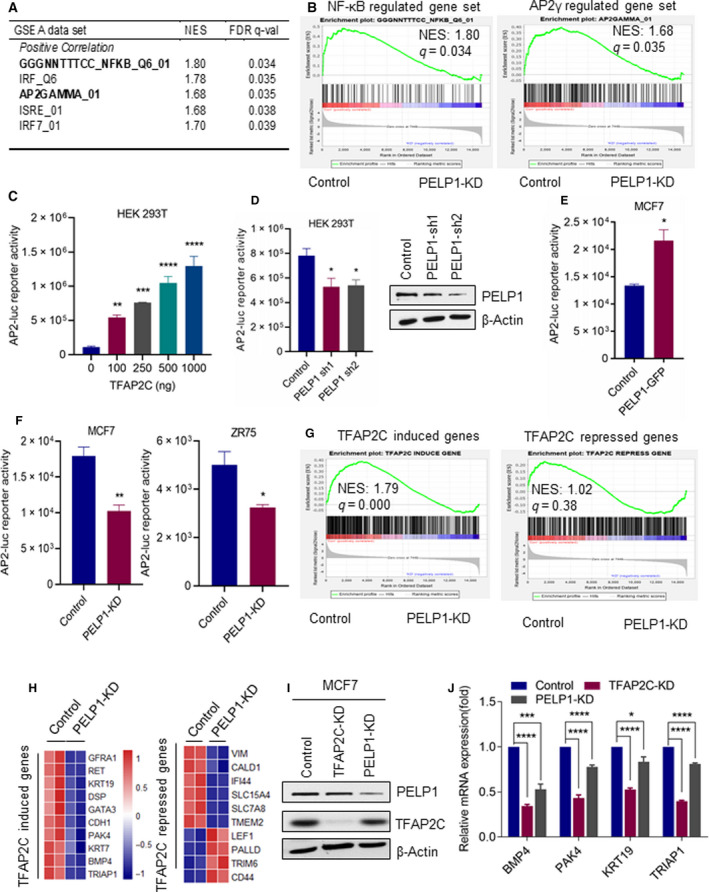
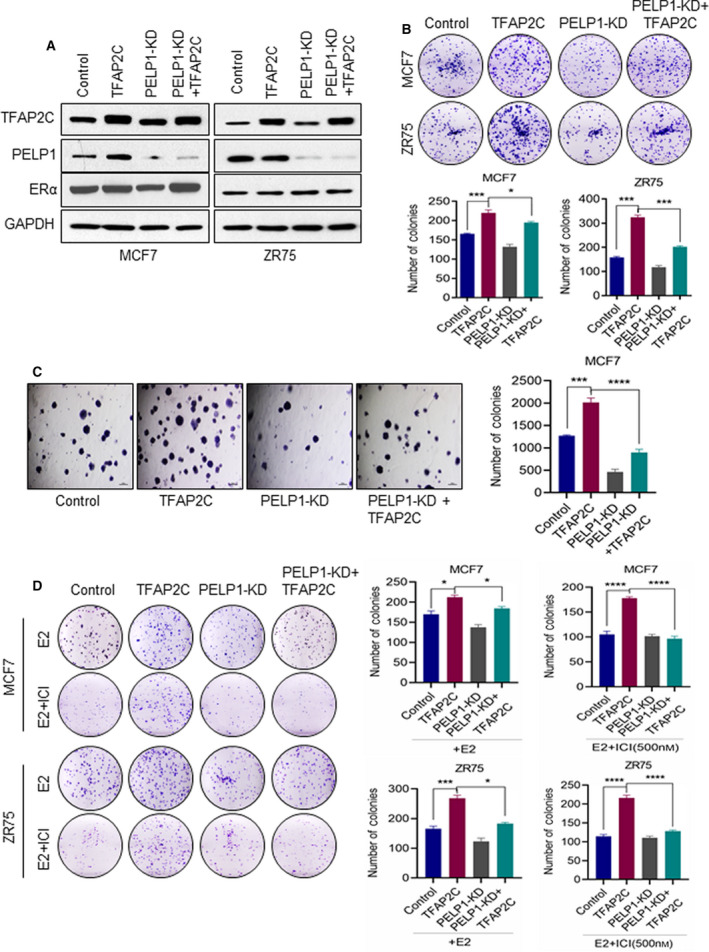
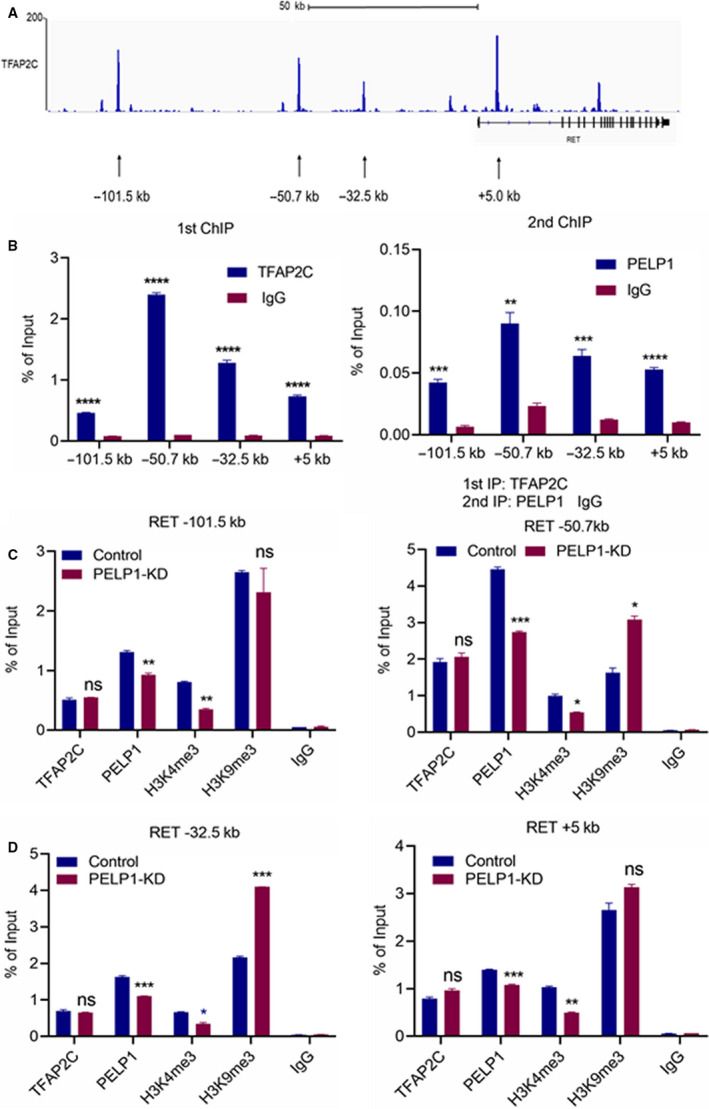
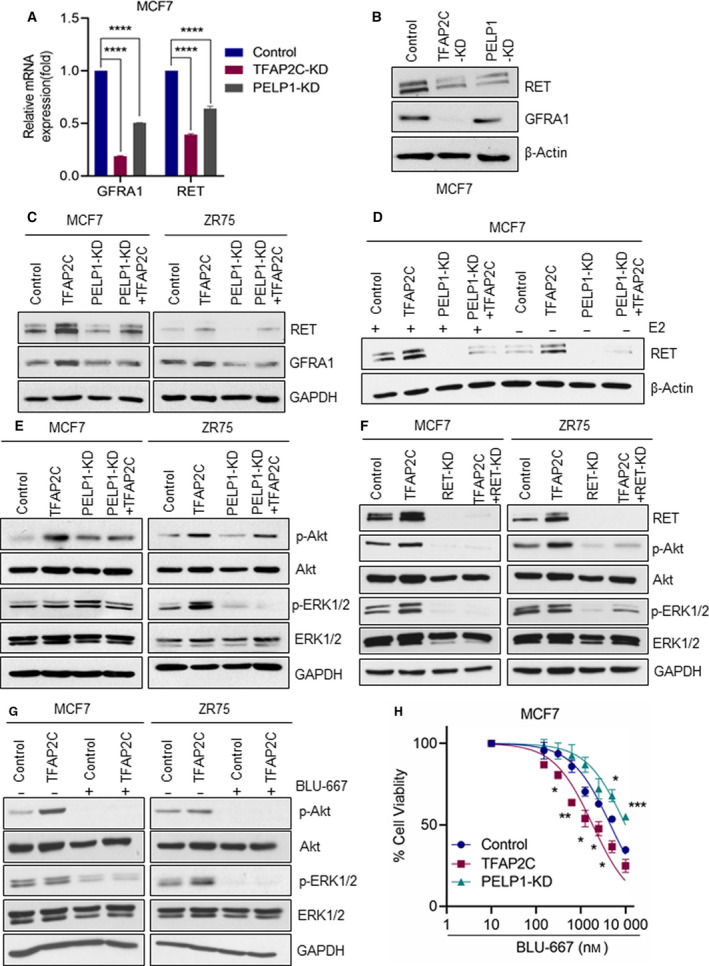
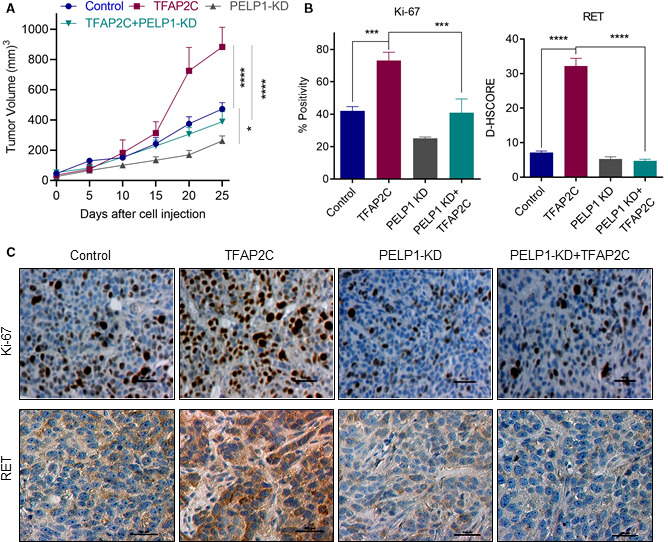
A significant proportion of estrogen receptor-positive (ER+) breast cancer (BC) initially responds to endocrine therapy but eventually evolves into therapy-resistant BC. Transcription factor AP-2 gamma (TFAP2C) is a known regulator of ER activity, and high expression of TFAP2C is associated with a decreased response to endocrine therapies. PELP1 is a nuclear receptor coregulator, commonly overexpressed in BC, and its levels are correlated with poorer survival. In this study, we identified PELP1 as a novel interacting protein of TFAP2C. RNA-seq analysis of PELP1 knockdown BC cells followed by transcription factor motif prediction pointed to TFAP2C being enriched in PELP1-regulated genes. Gene set enrichment analysis (GSEA) revealed that the TFAP2C-PELP1 axis induced a subset of common genes. Reporter gene assays confirmed PELP1 functions as a coactivator of TFAP2C. Mechanistic studies showed that PELP1-mediated changes in histone methylation contributed to increased expression of the TFAP2C target gene RET. Furthermore, the TFAP2C-PELP1 axis promoted the activation of the RET signaling pathway, which contributed to downstream activation of AKT and ERK pathways in ER+ BC cells. Concomitantly, knockdown of PELP1 attenuated these effects mediated by TFAP2C. Overexpression of TFAP2C contributed to increased cell proliferation and therapy resistance in ER+ BC models, while knockdown of PELP1 mitigated these effects. Utilizing ZR75-TFAP2C xenografts with or without PELP1 knockdown, we provided genetic evidence that endogenous PELP1 is essential for TFAP2C-driven BC progression in vivo. Collectively, our studies demonstrated that PELP1 plays a critical role in TFAP2C transcriptional and tumorigenic functions in BC and blocking the PELP1-TFAP2C axis could have utility for treating therapy resistance.
Keywords: PELP1; TFAP2C; breast cancer; coactivator; therapy resistance.
© 2020 The Authors. Molecular Oncology published by John Wiley & Sons Ltd on behalf of Federation of European BiochemicalSocieties.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Jager R, Werling U, Rimpf S, Jacob A & Schorle H (2003) Transcription factor AP‐2gamma stimulates proliferation and apoptosis and impairs differentiation in a transgenic model. Mol Cancer Res 1, 921–929. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
