

The cucumber mosaic virus 1a protein regulates interactions between the 2b protein and ARGONAUTE 1 while maintaining the silencing suppressor activity of the 2b protein
- PMID: 33270799
- PMCID: PMC7738167
- DOI: 10.1371/journal.ppat.1009125
The cucumber mosaic virus 1a protein regulates interactions between the 2b protein and ARGONAUTE 1 while maintaining the silencing suppressor activity of the 2b protein
Abstract
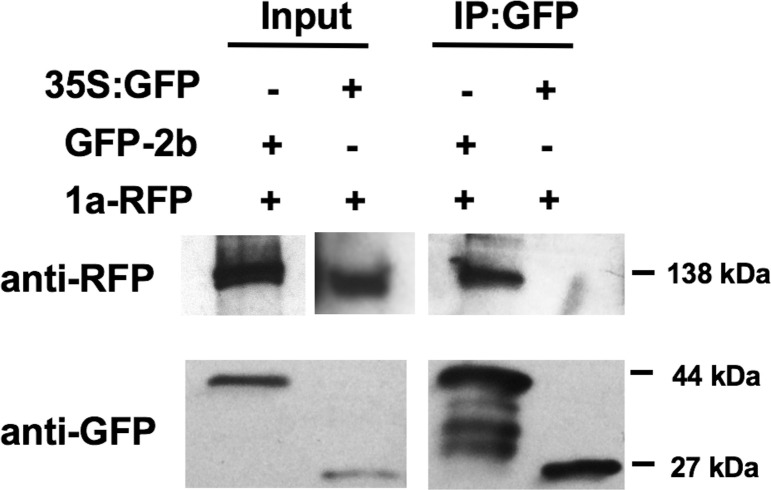
The cucumber mosaic virus (CMV) 2b viral suppressor of RNA silencing (VSR) is a potent counter-defense and pathogenicity factor that inhibits antiviral silencing by titration of short double-stranded RNAs. It also disrupts microRNA-mediated regulation of host gene expression by binding ARGONAUTE 1 (AGO1). But in Arabidopsis thaliana complete inhibition of AGO1 is counterproductive to CMV since this triggers another layer of antiviral silencing mediated by AGO2, de-represses strong resistance against aphids (the insect vectors of CMV), and exacerbates symptoms. Using confocal laser scanning microscopy, bimolecular fluorescence complementation, and co-immunoprecipitation assays we found that the CMV 1a protein, a component of the viral replicase complex, regulates the 2b-AGO1 interaction. By binding 2b protein molecules and sequestering them in P-bodies, the 1a protein limits the proportion of 2b protein molecules available to bind AGO1, which ameliorates 2b-induced disease symptoms, and moderates induction of resistance to CMV and to its aphid vector. However, the 1a protein-2b protein interaction does not inhibit the ability of the 2b protein to inhibit silencing of reporter gene expression in agroinfiltration assays. The interaction between the CMV 1a and 2b proteins represents a novel regulatory system in which specific functions of a VSR are selectively modulated by another viral protein. The finding also provides a mechanism that explains how CMV, and possibly other viruses, modulates symptom induction and manipulates host-vector interactions.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures









References
-
- Yoon JY, Palukaitis P, Choi SK (2019) Host Range In: Palukaitis P, García-Arenal F, editors. Cucumber Mosaic Virus. St Paul MN: APS Press; pp. 15–18.
-
- Jacquemond M (2012) Cucumber mosaic virus. Adv Virus Res 84: 439–504 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
