

A dual role for α-synuclein in facilitation and depression of dopamine release from substantia nigra neurons in vivo
- PMID: 33273122
- PMCID: PMC7768743
- DOI: 10.1073/pnas.2013652117
A dual role for α-synuclein in facilitation and depression of dopamine release from substantia nigra neurons in vivo
Abstract
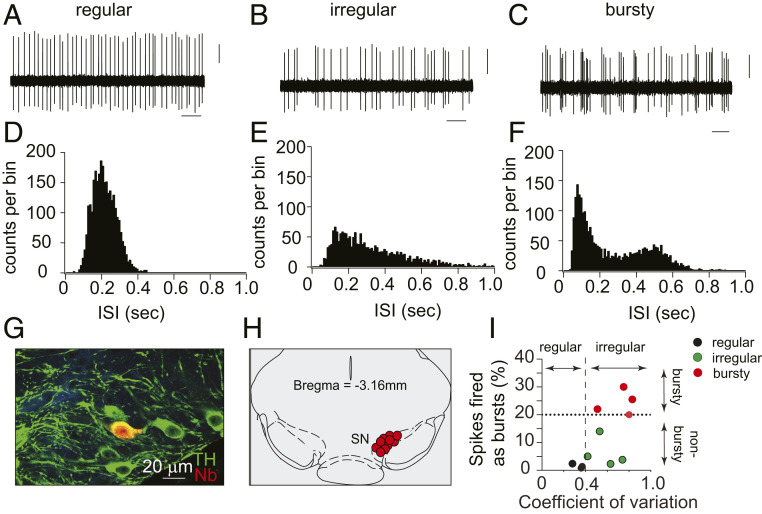
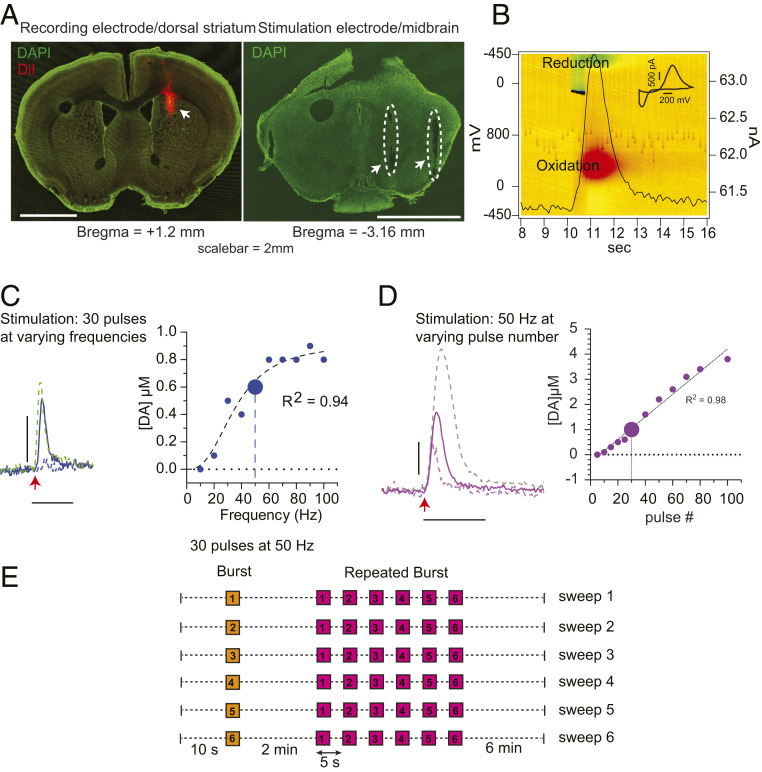
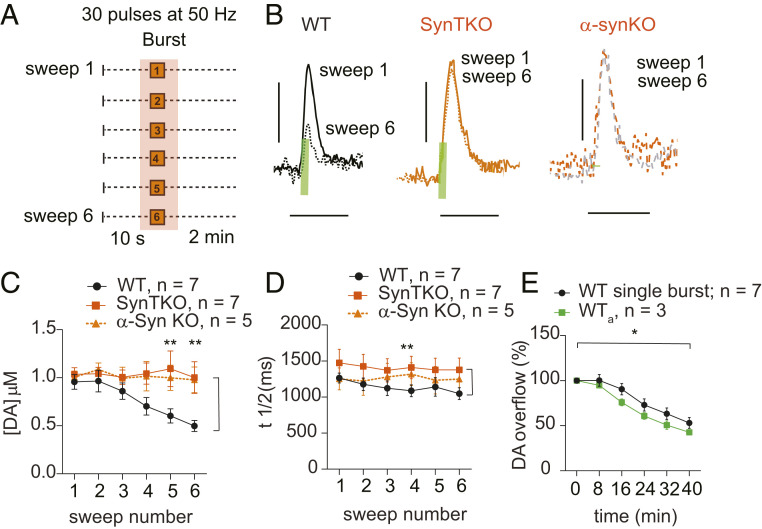
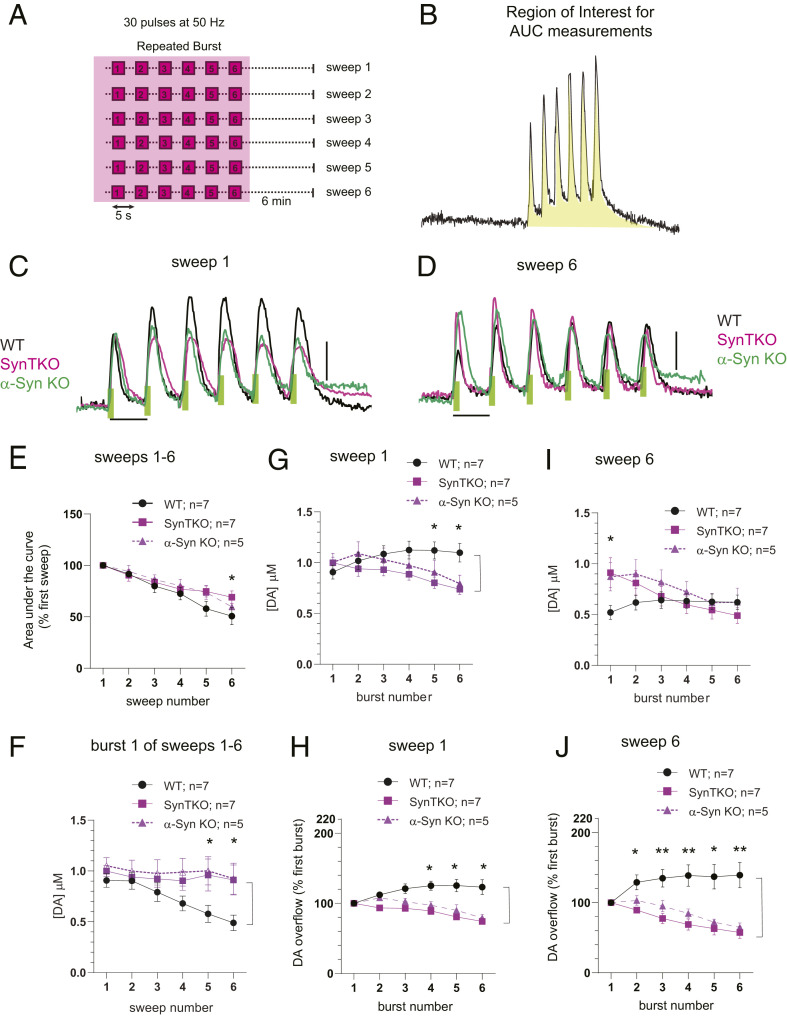
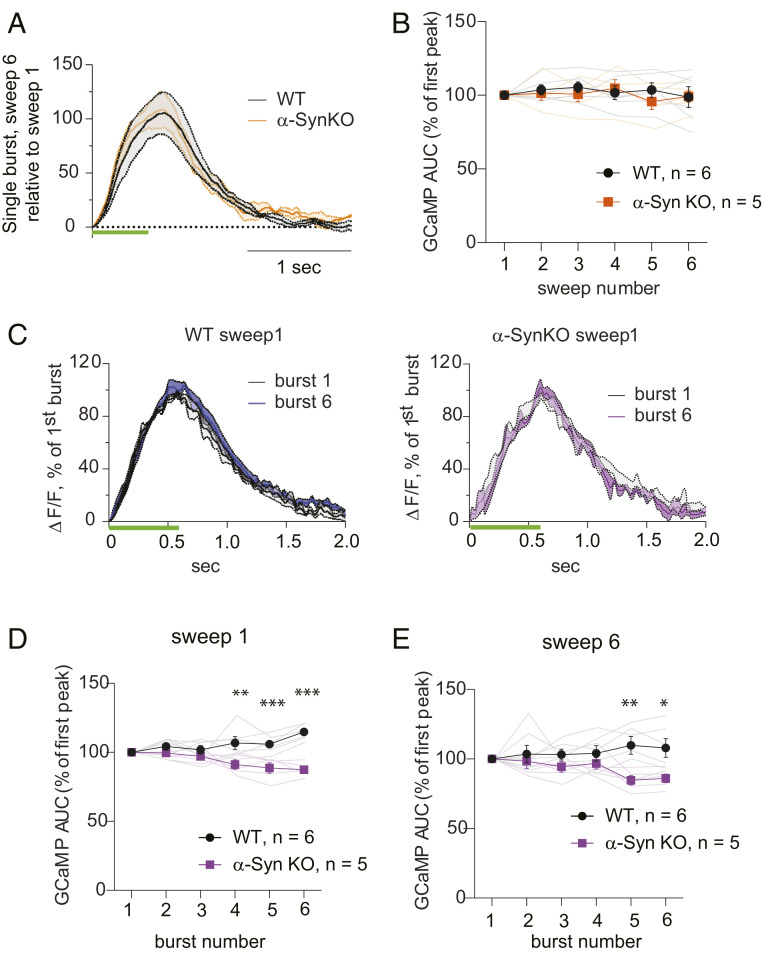
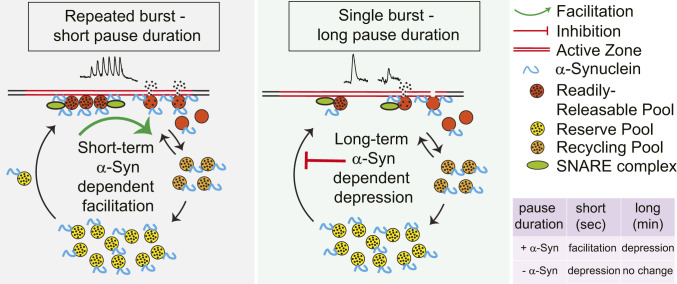
α-Synuclein is expressed at high levels at presynaptic terminals, but defining its role in the regulation of neurotransmission under physiologically relevant conditions has proven elusive. We report that, in vivo, α-synuclein is responsible for the facilitation of dopamine release triggered by action potential bursts separated by short intervals (seconds) and a depression of release with longer intervals between bursts (minutes). These forms of presynaptic plasticity appear to be independent of the presence of β- and γ-synucleins or effects on presynaptic calcium and are consistent with a role for synucleins in the enhancement of synaptic vesicle fusion and turnover. These results indicate that the presynaptic effects of α-synuclein depend on specific patterns of neuronal activity.
Keywords: alpha-Synuclein; dopamine; in vivo neurotransmission.
Copyright © 2020 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Nakajo S., et al. , Purification and characterization of a novel brain-specific 14-kDa protein. J. Neurochem. 55, 2031–2038 (1990). - PubMed
-
- Iwai A., et al. , The precursor protein of non-A beta component of Alzheimer’s disease amyloid is a presynaptic protein of the central nervous system. Neuron 14, 467–475 (1995). - PubMed
-
- Jakes R., Spillantini M. G., Goedert M., Identification of two distinct synucleins from human brain. FEBS Lett. 345, 27–32 (1994). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
