

Ubiquitination of phytoene synthase 1 precursor modulates carotenoid biosynthesis in tomato
- PMID: 33273697
- PMCID: PMC7713427
- DOI: 10.1038/s42003-020-01474-3
Ubiquitination of phytoene synthase 1 precursor modulates carotenoid biosynthesis in tomato
Abstract
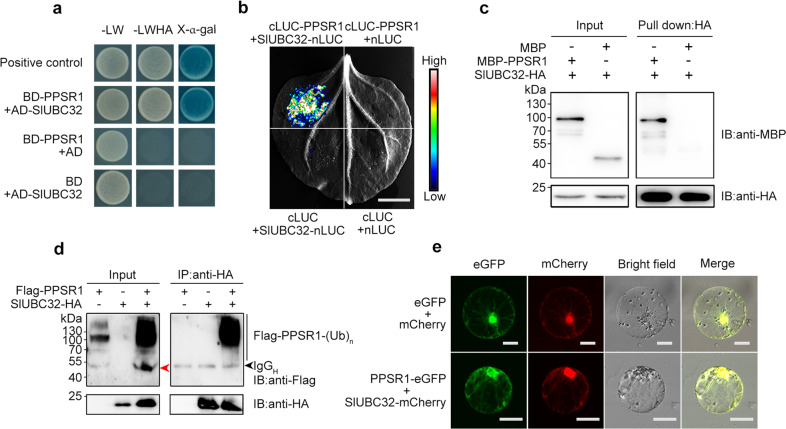
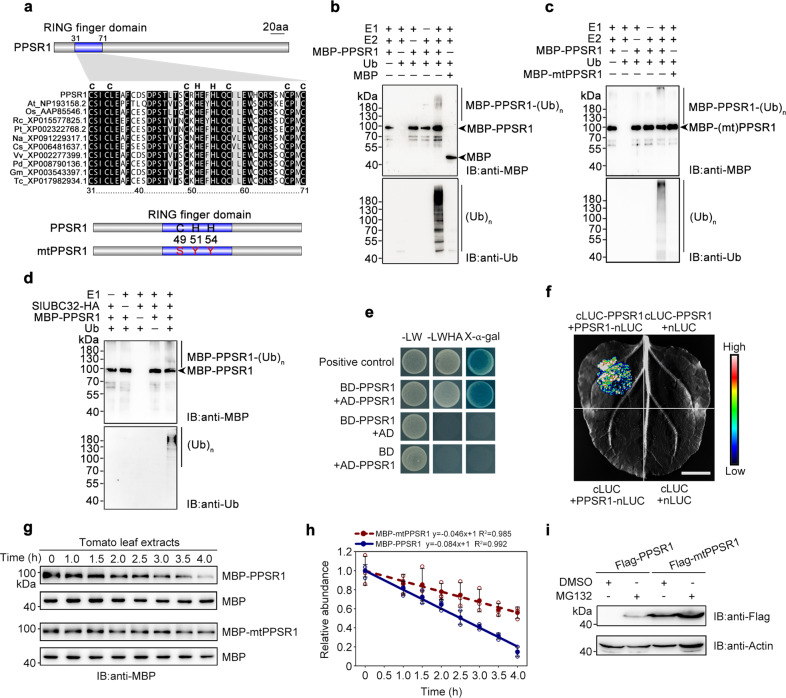
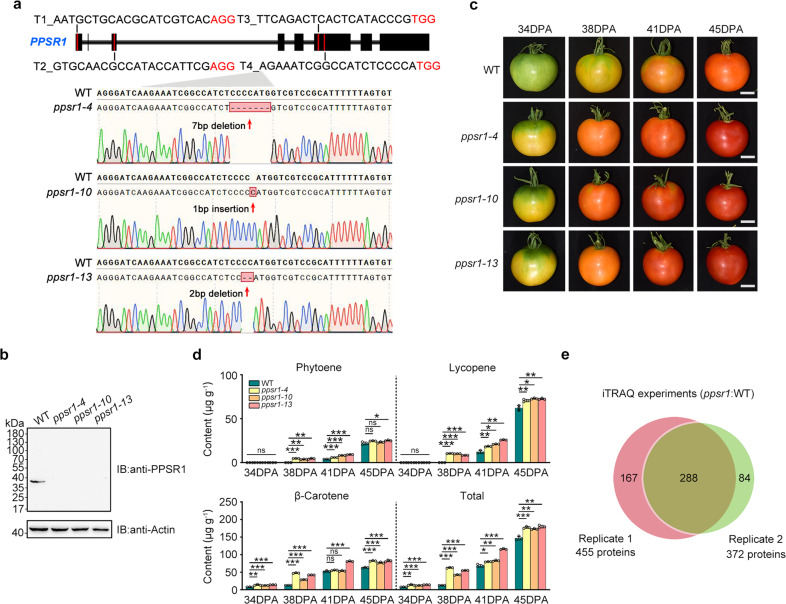
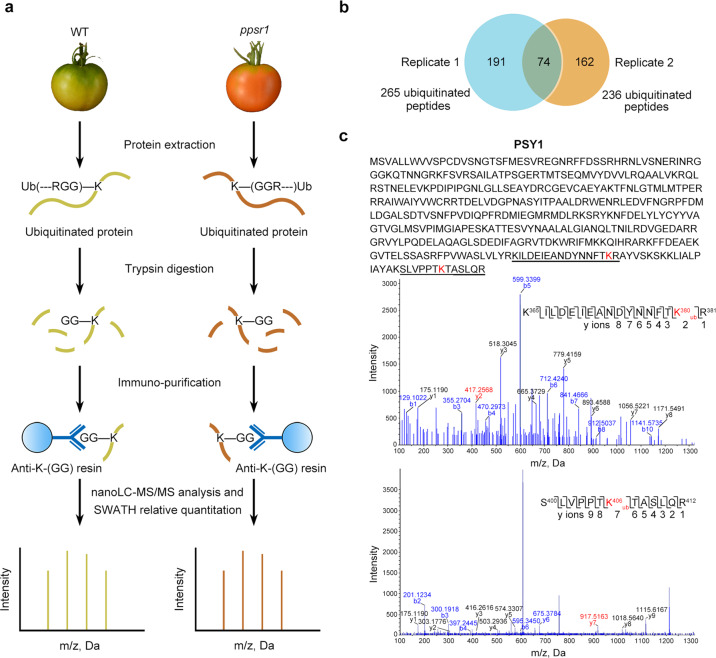
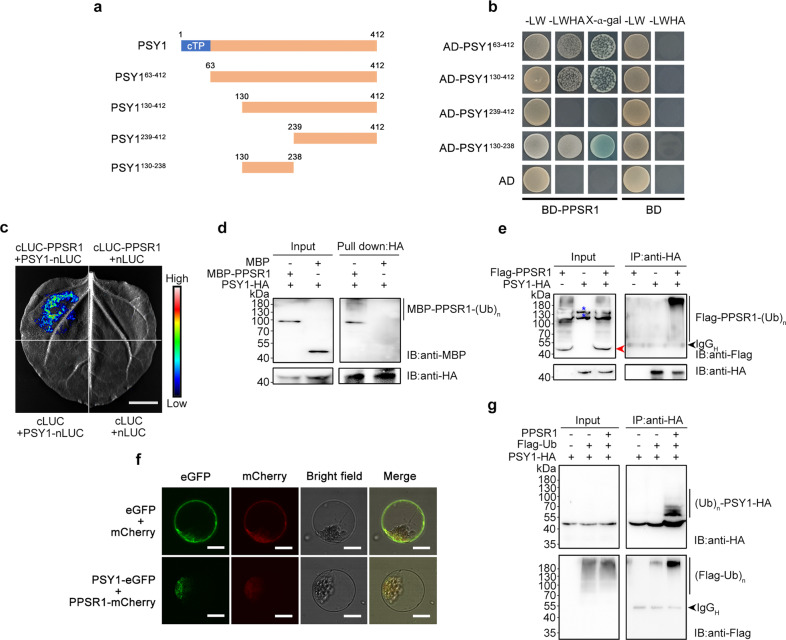
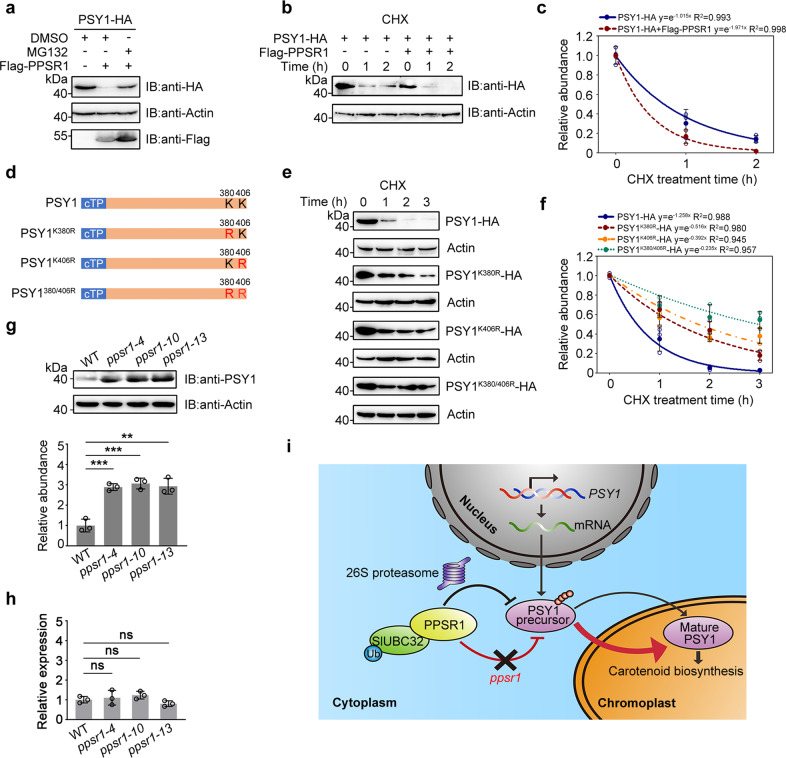
Carotenoids are natural pigments that are indispensable to plants and humans, whereas the regulation of carotenoid biosynthesis by post-translational modification remains elusive. Here, we show that a tomato E3 ubiquitin ligase, Plastid Protein Sensing RING E3 ligase 1 (PPSR1), is responsible for the regulation of carotenoid biosynthesis. PPSR1 exhibits self-ubiquitination activity and loss of PPSR1 function leads to an increase in carotenoids in tomato fruit. PPSR1 affects the abundance of 288 proteins, including phytoene synthase 1 (PSY1), the key rate-limiting enzyme in the carotenoid biosynthetic pathway. PSY1 contains two ubiquitinated lysine residues (Lys380 and Lys406) as revealed by the global analysis and characterization of protein ubiquitination. We provide evidence that PPSR1 interacts with PSY1 precursor protein and mediates its degradation via ubiquitination, thereby affecting the steady-state level of PSY1 protein. Our findings not only uncover a regulatory mechanism for controlling carotenoid biosynthesis, but also provide a strategy for developing carotenoid-enriched horticultural crops.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
