

Phosphorylation of RIAM by src promotes integrin activation by unmasking the PH domain of RIAM
- PMID: 33275877
- PMCID: PMC8026550
- DOI: 10.1016/j.str.2020.11.011
Phosphorylation of RIAM by src promotes integrin activation by unmasking the PH domain of RIAM
Abstract
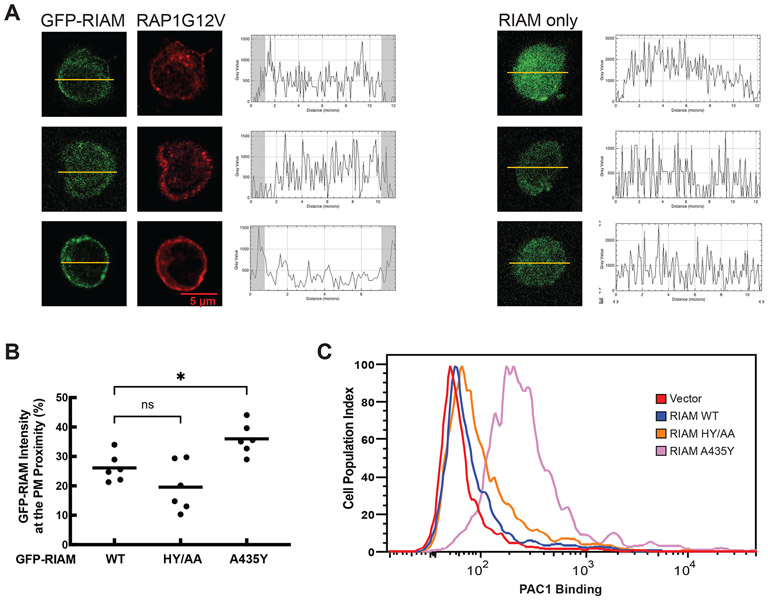
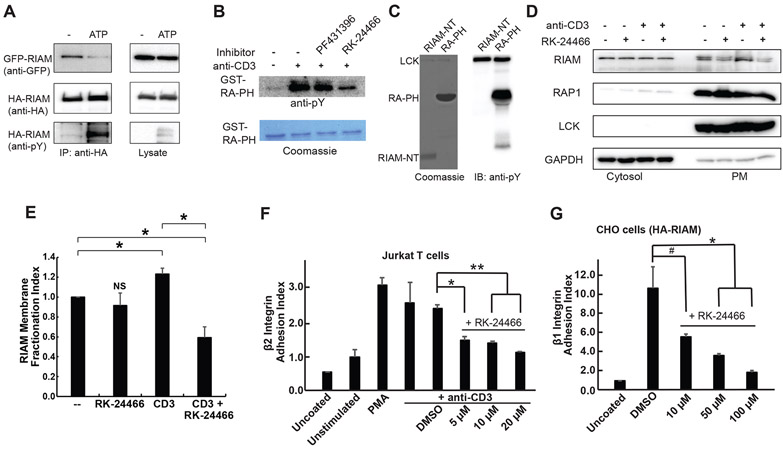
Integrin activation controls cell adhesion, migration, invasion, and extracellular matrix remodeling. RIAM (RAP1-GTP-interacting adaptor molecule) is recruited by activated RAP1 to the plasma membrane (PM) to mediate integrin activation via an inside-out signaling pathway. This process requires the association of the pleckstrin homology (PH) domain of RIAM with the membrane PIP2. We identify a conserved intermolecular interface that masks the PIP2-binding site in the PH domains of RIAM. Our data indicate that phosphorylation of RIAM by Src family kinases disrupts this PH-mediated interface, unmasks the membrane PIP2-binding site, and promotes integrin activation. We further demonstrate that this process requires phosphorylation of Tyr267 and Tyr427 in the RIAM PH domain by Src. Our data reveal an unorthodox regulatory mechanism of small GTPase effector proteins by phosphorylation-dependent PM association of the PH domain and provide new insights into the link between Src kinases and integrin signaling.
Keywords: FYN; LCK; PH domain; PIP2 binding; RAP1; RIAM; Src kinase; integrin signaling; lamellipodin; phosphorylation.
Copyright © 2020 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures



Comment in
-
Src-mediated phosphorylation of RIAM promotes integrin activation.Structure. 2021 Apr 1;29(4):305-307. doi: 10.1016/j.str.2021.03.009. Structure. 2021. PMID: 33798425
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
