

Mapping the Degradable Kinome Provides a Resource for Expedited Degrader Development
- PMID: 33275901
- PMCID: PMC10294644
- DOI: 10.1016/j.cell.2020.10.038
Mapping the Degradable Kinome Provides a Resource for Expedited Degrader Development
Abstract
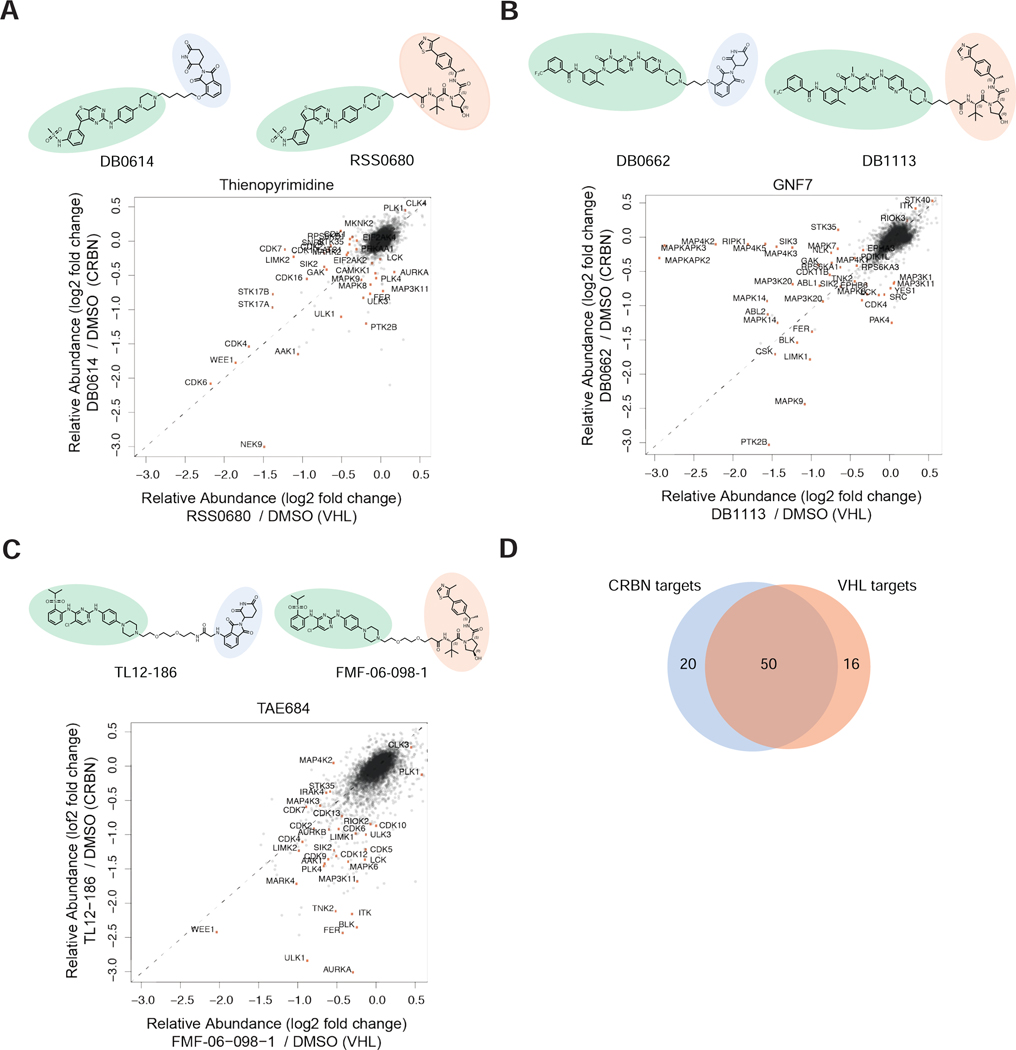
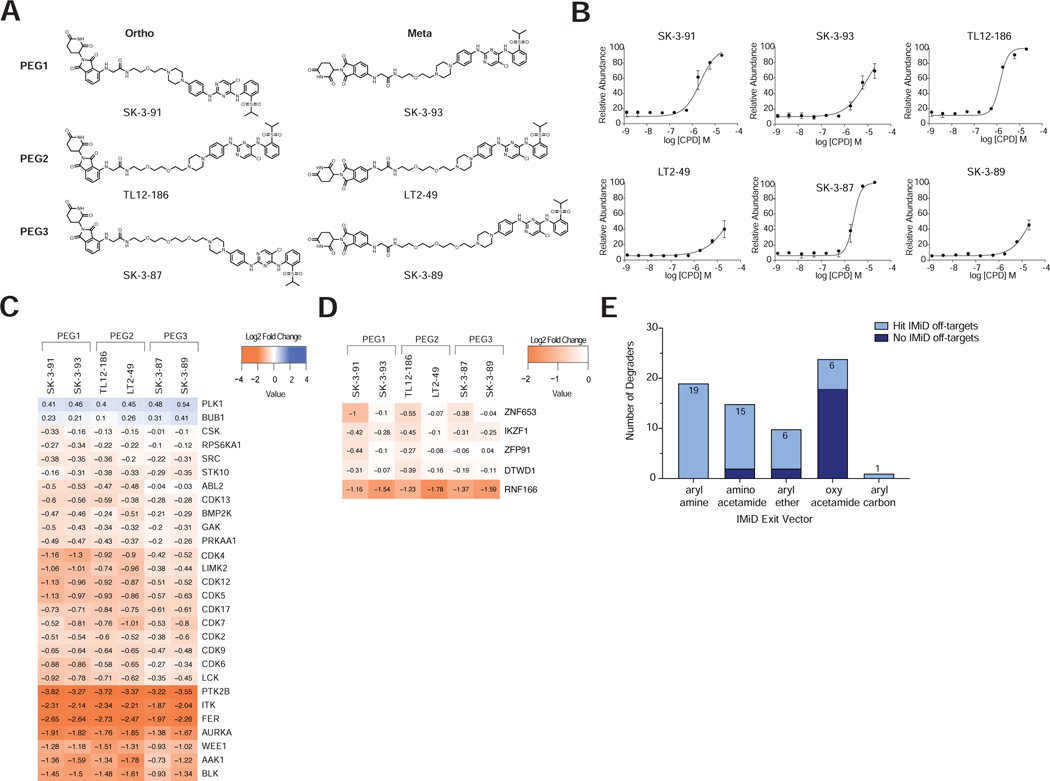
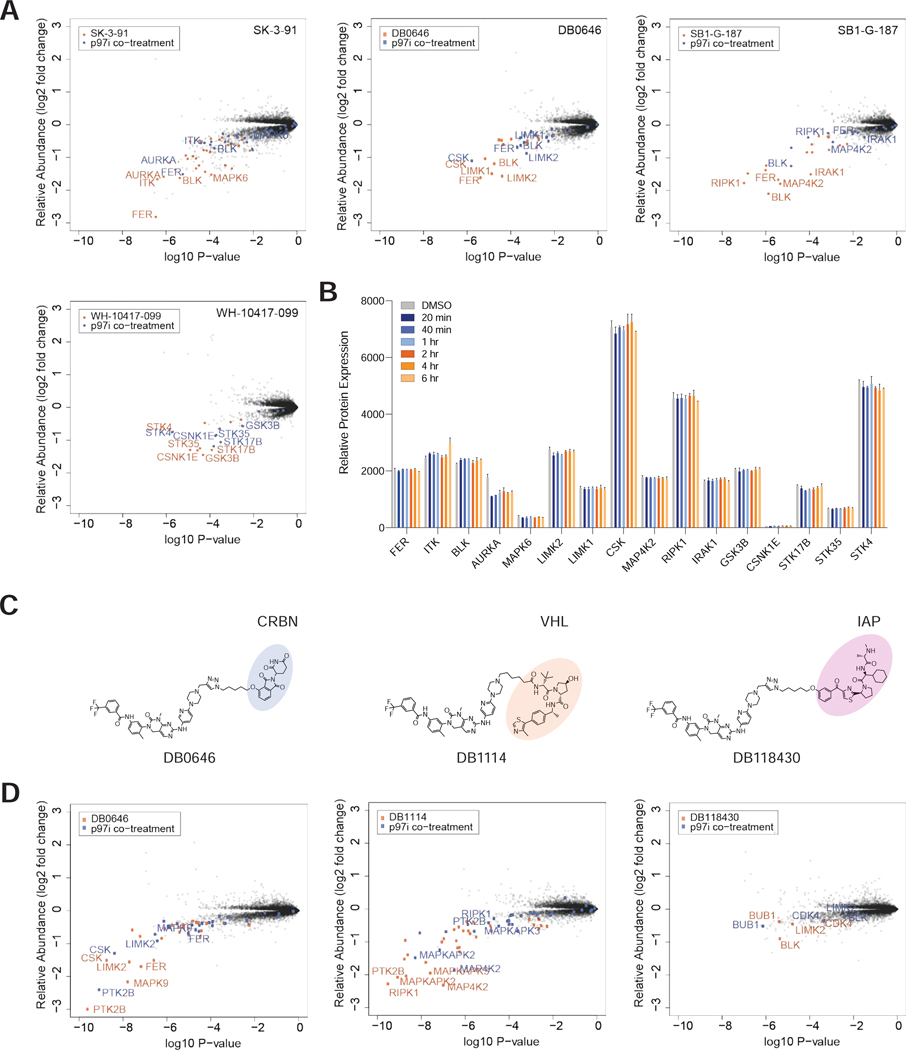
Targeted protein degradation (TPD) refers to the use of small molecules to induce ubiquitin-dependent degradation of proteins. TPD is of interest in drug development, as it can address previously inaccessible targets. However, degrader discovery and optimization remains an inefficient process due to a lack of understanding of the relative importance of the key molecular events required to induce target degradation. Here, we use chemo-proteomics to annotate the degradable kinome. Our expansive dataset provides chemical leads for ∼200 kinases and demonstrates that the current practice of starting from the highest potency binder is an ineffective method for discovering active compounds. We develop multitargeted degraders to answer fundamental questions about the ubiquitin proteasome system, uncovering that kinase degradation is p97 dependent. This work will not only fuel kinase degrader discovery, but also provides a blueprint for evaluating targeted degradation across entire gene families to accelerate understanding of TPD beyond the kinome.
Keywords: E3 ligase; IMiD; PROTAC; degrader; kinase; targeted degradation; ubiquitin; ubiquitin proteasome system.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests N.S.G. is a founder, science advisory board (SAB) member, and equity holder in Gatekeeper, Syros, Petra, C4, B2S, Aduro, Jengu, and Soltego (board member). The Gray lab receives or has received research funding from Novartis, Takeda, Astellas, Taiho, Janssen, Kinogen, Voronoi, Her2llc, Deerfield, and Sanofi. E.S.F. is a founder, science advisory board member, and equity holder in Civetta, Jengu (board member), and Neomorph, an equity holder in C4, and a consultant to Astellas, Novartis, Deerfield, and EcoR1. The Fischer lab receives or has received research funding from Novartis, Astellas, and Deerfield. K.A.D., F.M.F., J.W.B., T.S., E.S.F., and N.S.G. are inventors on a patent application relating to this work, owned by DFCI.
Figures
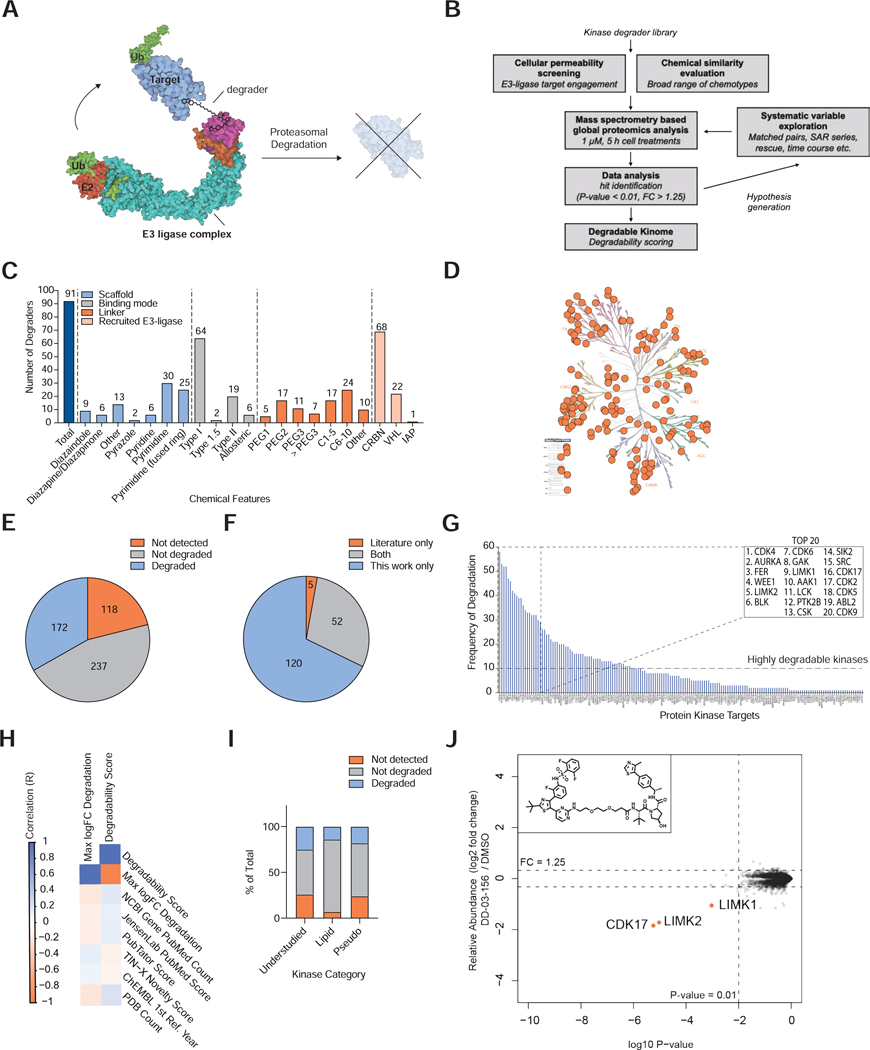
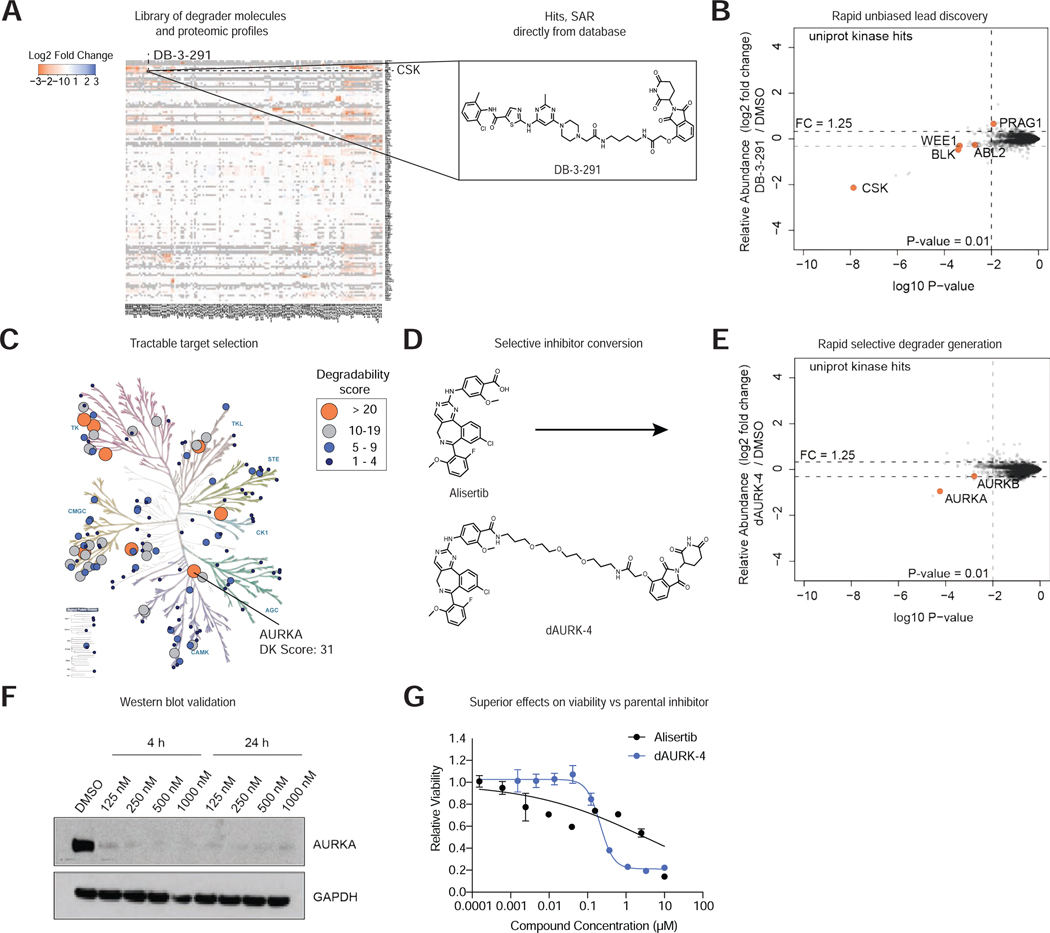
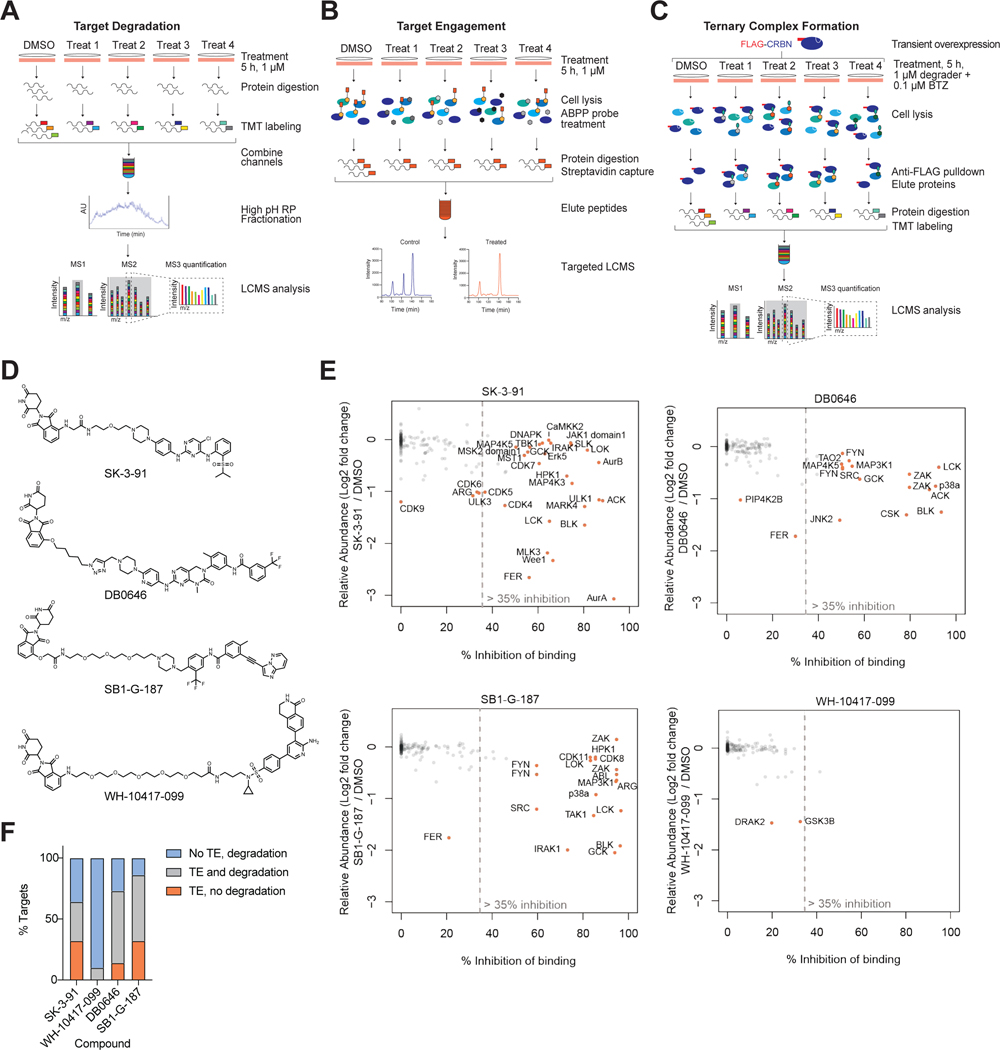
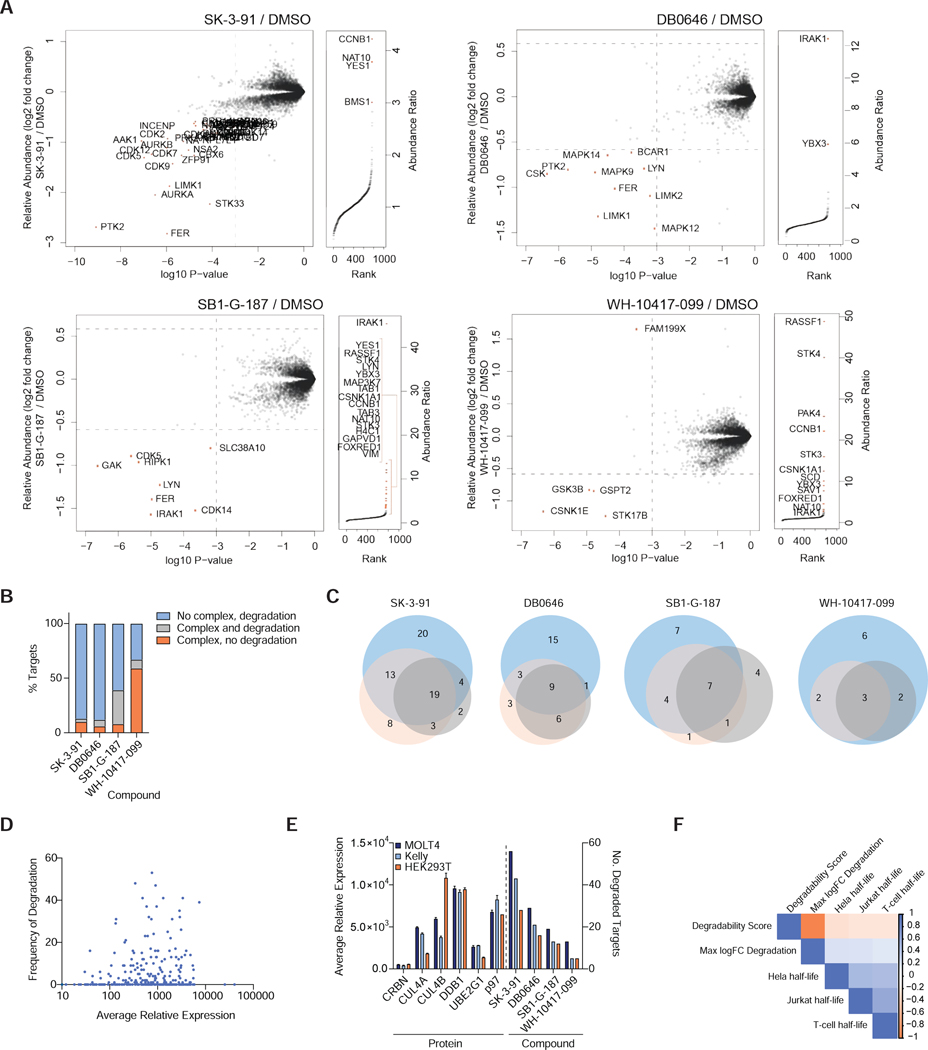
References
-
- Bingqi T, Mai L, Yi X, Jessica S, John A, Jeffrey M,M, Markus S, Thomas J,M, and Daniel N. (2020). Targeted Protein Degradation via a Covalent Reversible Degrader Based on Bardoxolone. ChemRxiv.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
