

The substrate specificity of the human TRAPPII complex's Rab-guanine nucleotide exchange factor activity
- PMID: 33277614
- PMCID: PMC7719173
- DOI: 10.1038/s42003-020-01459-2
The substrate specificity of the human TRAPPII complex's Rab-guanine nucleotide exchange factor activity
Abstract
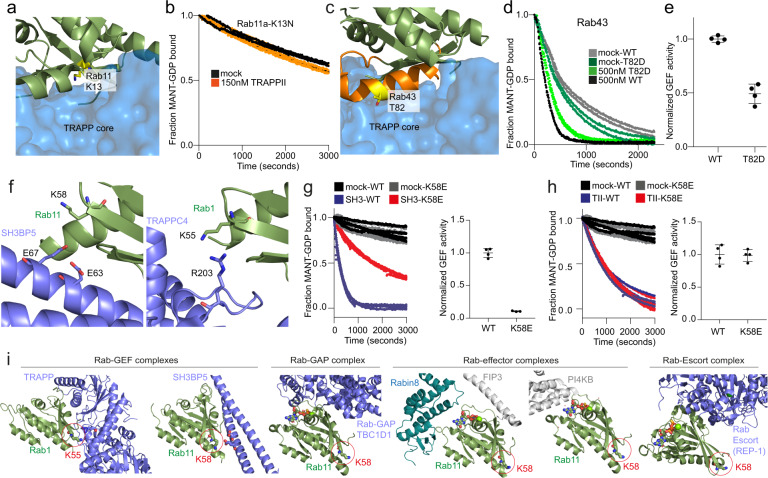
The TRAnsport Protein Particle (TRAPP) complexes act as Guanine nucleotide exchange factors (GEFs) for Rab GTPases, which are master regulators of membrane trafficking in eukaryotic cells. In metazoans, there are two large multi-protein TRAPP complexes: TRAPPII and TRAPPIII, with the TRAPPII complex able to activate both Rab1 and Rab11. Here we present detailed biochemical characterisation of Rab-GEF specificity of the human TRAPPII complex, and molecular insight into Rab binding. GEF assays of the TRAPPII complex against a panel of 20 different Rab GTPases revealed GEF activity on Rab43 and Rab19. Electron microscopy and chemical cross-linking revealed the architecture of mammalian TRAPPII. Hydrogen deuterium exchange MS showed that Rab1, Rab11 and Rab43 share a conserved binding interface. Clinical mutations in Rab11, and phosphomimics of Rab43, showed decreased TRAPPII GEF mediated exchange. Finally, we designed a Rab11 mutation that maintained TRAPPII-mediated GEF activity while decreasing activity of the Rab11-GEF SH3BP5, providing a tool to dissect Rab11 signalling. Overall, our results provide insight into the GTPase specificity of TRAPPII, and how clinical mutations disrupt this regulation.
Conflict of interest statement
The authors declare no competing interests.
Figures
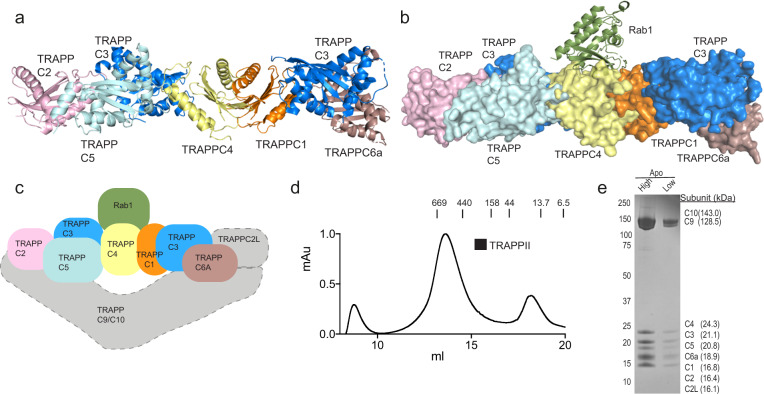
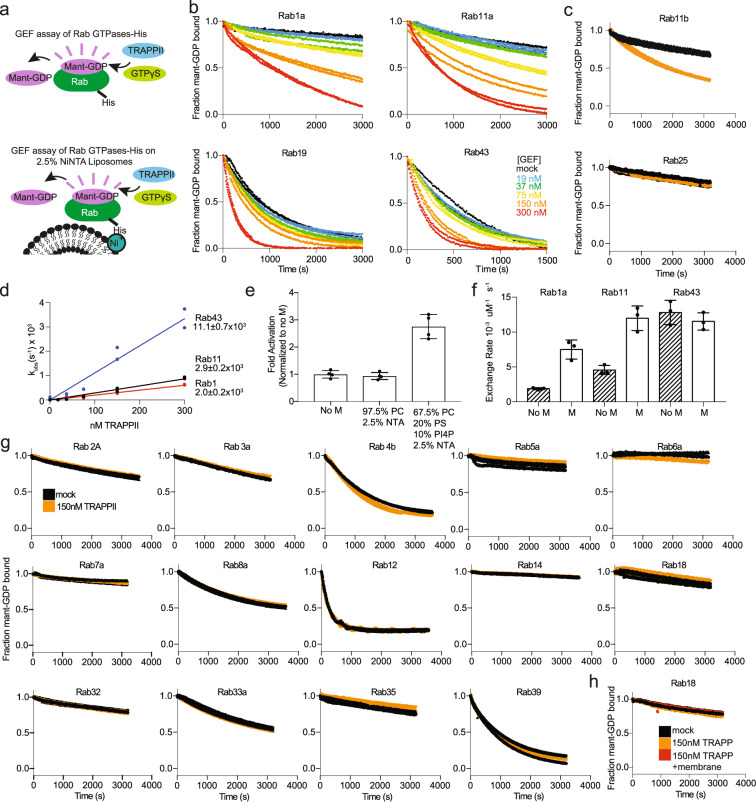
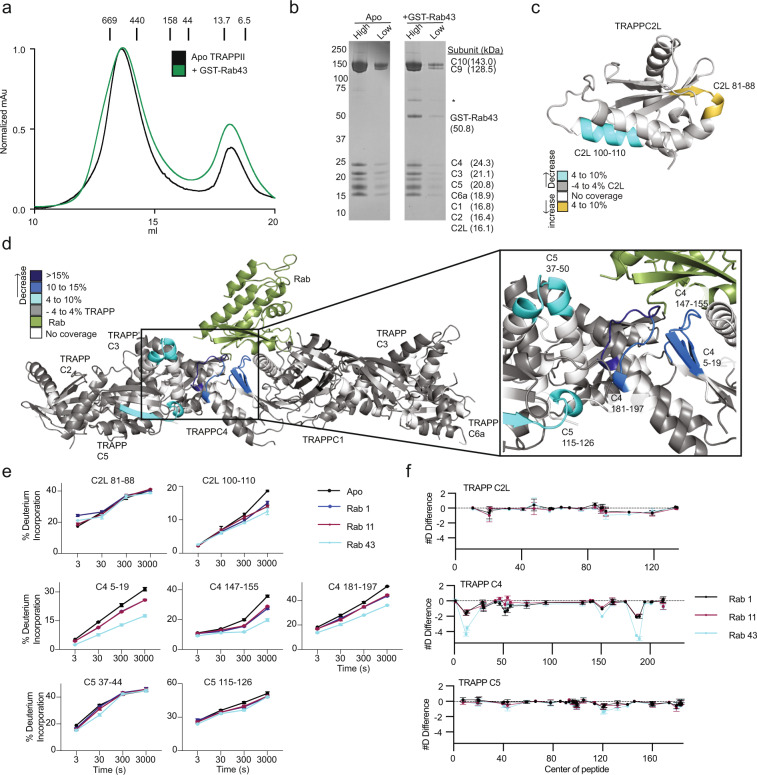
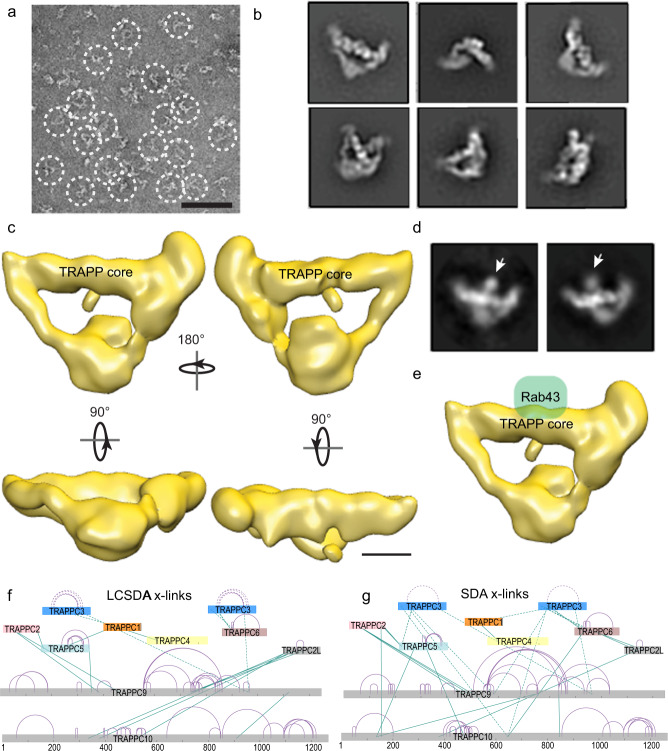
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
