

Iron: An Essential Element of Cancer Metabolism
- PMID: 33287315
- PMCID: PMC7761773
- DOI: 10.3390/cells9122591
Iron: An Essential Element of Cancer Metabolism
Abstract
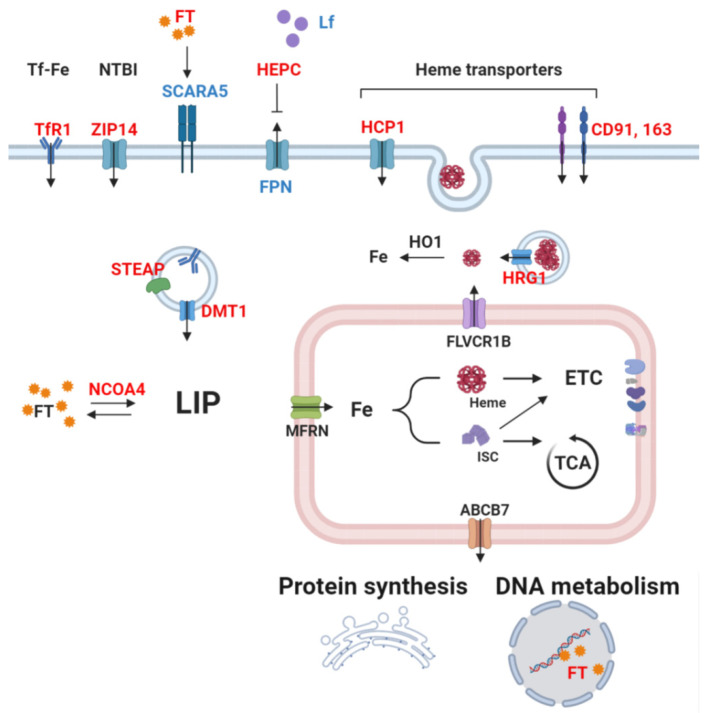
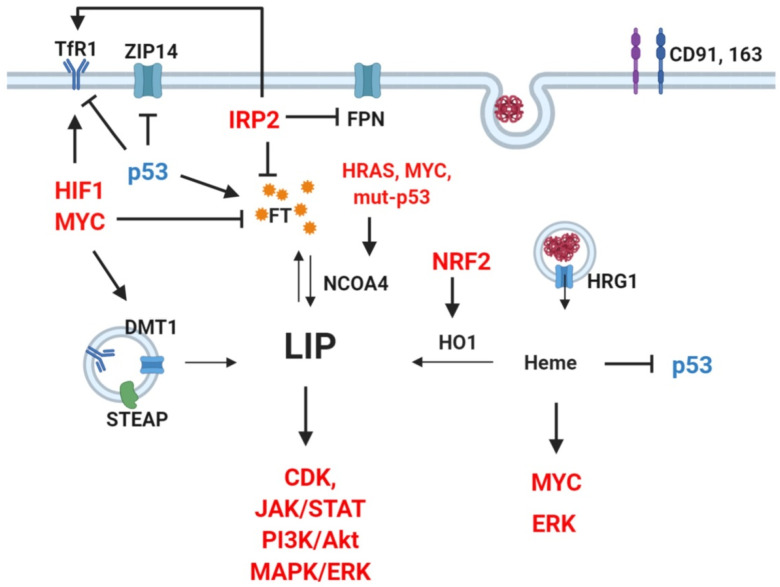
Cancer cells undergo considerable metabolic changes to foster uncontrolled proliferation in a hostile environment characterized by nutrient deprivation, poor vascularization and immune infiltration. While metabolic reprogramming has been recognized as a hallmark of cancer, the role of micronutrients in shaping these adaptations remains scarcely investigated. In particular, the broad electron-transferring abilities of iron make it a versatile cofactor that is involved in a myriad of biochemical reactions vital to cellular homeostasis, including cell respiration and DNA replication. In cancer patients, systemic iron metabolism is commonly altered. Moreover, cancer cells deploy diverse mechanisms to increase iron bioavailability to fuel tumor growth. Although iron itself can readily participate in redox reactions enabling vital processes, its reactivity also gives rise to reactive oxygen species (ROS). Hence, cancer cells further rely on antioxidant mechanisms to withstand such stress. The present review provides an overview of the common alterations of iron metabolism occurring in cancer and the mechanisms through which iron promotes tumor growth.
Keywords: cancer metabolism; iron; iron-sulfur cluster; mitochondria.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
