

Analysis of Complex DNA Rearrangements during Early Stages of HAC Formation
- PMID: 33289546
- PMCID: PMC7754191
- DOI: 10.1021/acssynbio.0c00326
Analysis of Complex DNA Rearrangements during Early Stages of HAC Formation
Abstract
Human artificial chromosomes (HACs) are important tools for epigenetic engineering, for measuring chromosome instability (CIN), and for possible gene therapy. However, their use in the latter is potentially limited because the input HAC-seeding DNA can undergo an unpredictable series of rearrangements during HAC formation. As a result, after transfection and HAC formation, each cell clone contains a HAC with a unique structure that cannot be precisely predicted from the structure of the HAC-seeding DNA. Although it has been reported that these rearrangements can happen, the timing and mechanism of their formation has yet to be described. Here we synthesized a HAC-seeding DNA with two distinct structural domains and introduced it into HT1080 cells. We characterized a number of HAC-containing clones and subclones to track DNA rearrangements during HAC establishment. We demonstrated that rearrangements can occur early during HAC formation. Subsequently, the established HAC genomic organization is stably maintained across many cell generations. Thus, early stages in HAC formation appear to at least occasionally involve a process of DNA shredding and shuffling that resembles chromothripsis, an important hallmark of many cancer types. Understanding these events during HAC formation has critical implications for future efforts aimed at synthesizing and exploiting synthetic human chromosomes.
Keywords: CENP-A; centromere; epigenetic engineering; human artificial chromosome; kinetochore; mitosis.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
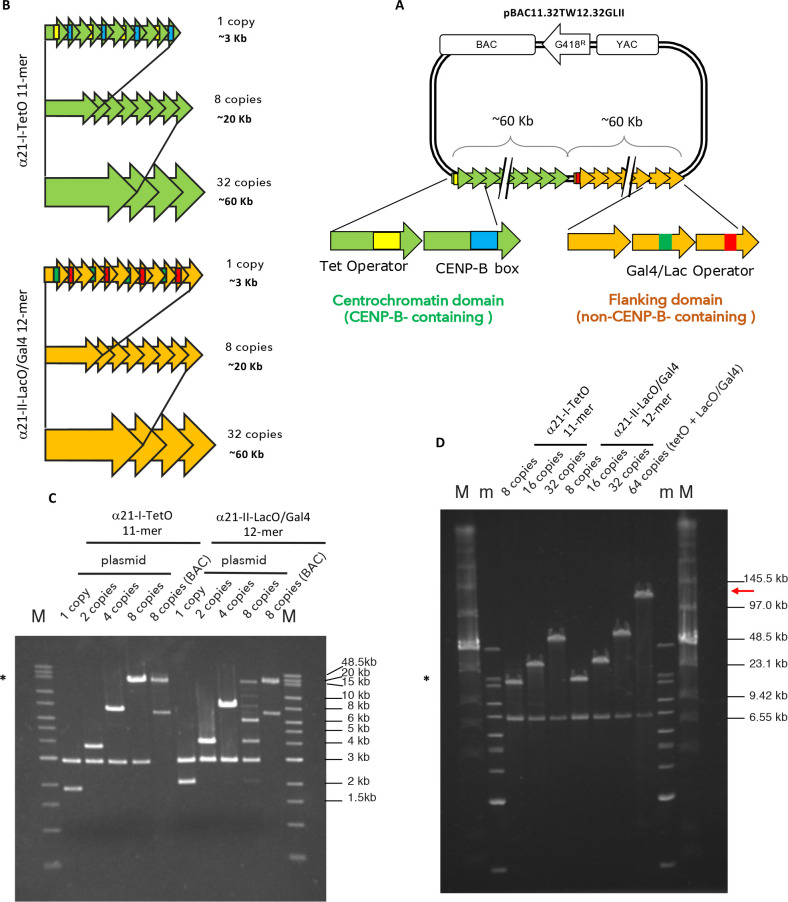
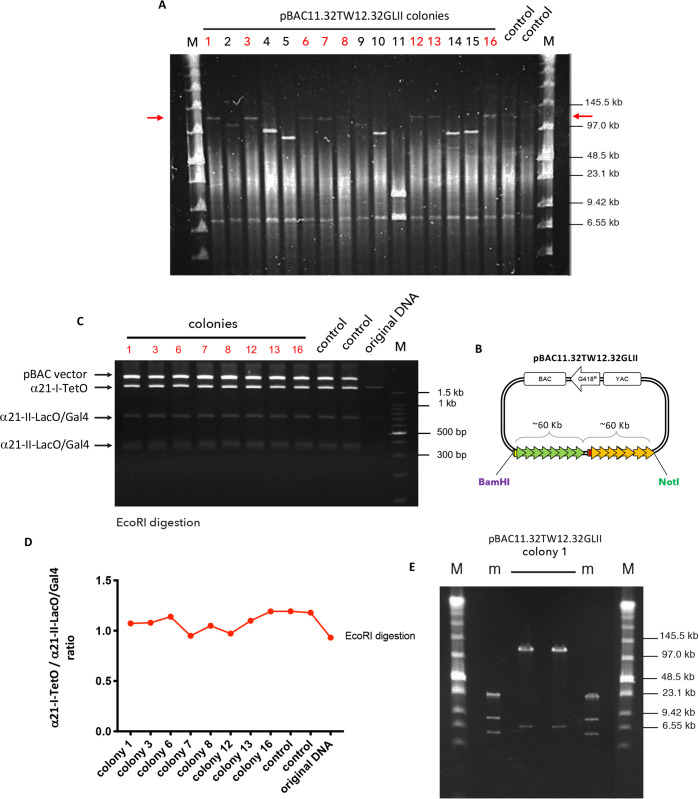
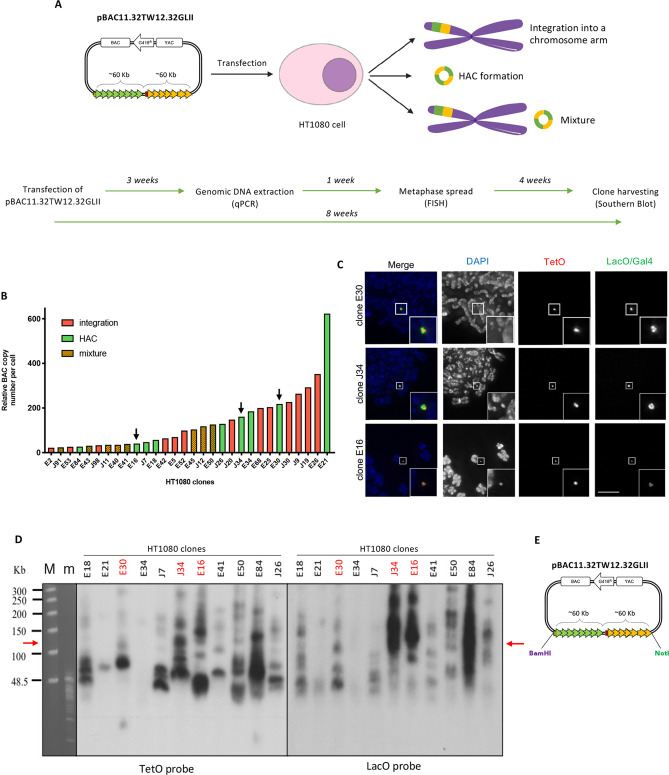
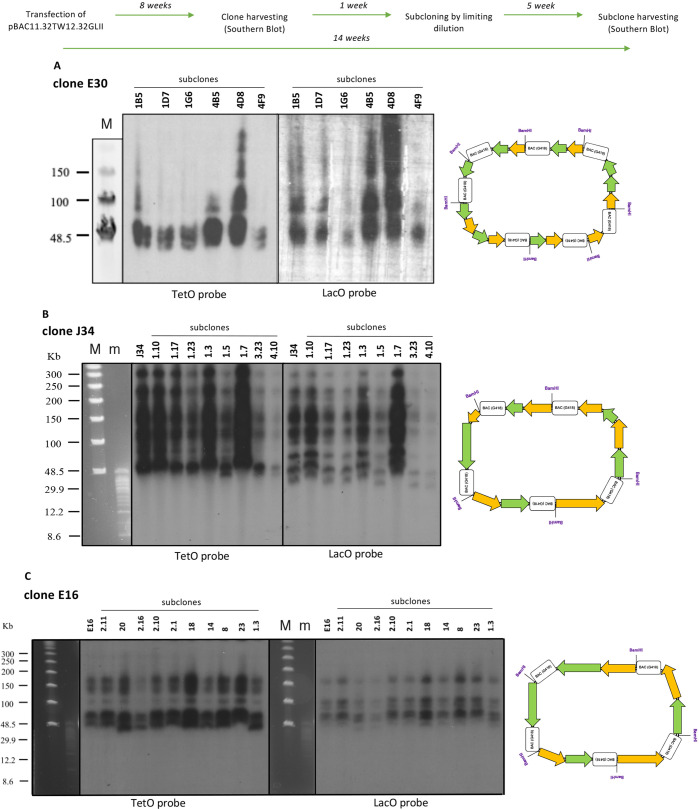
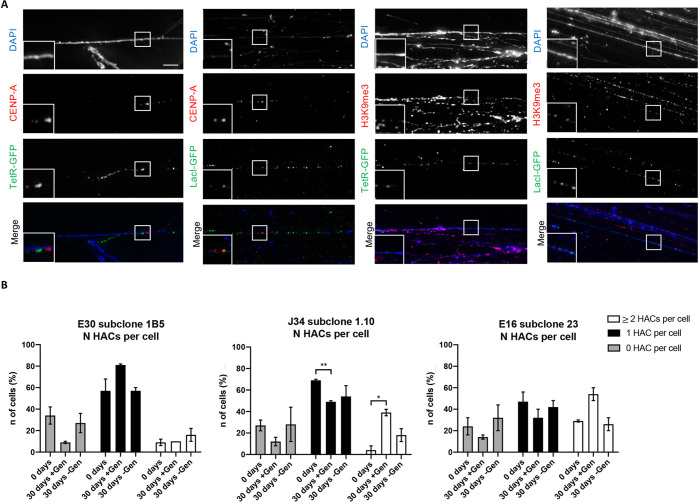
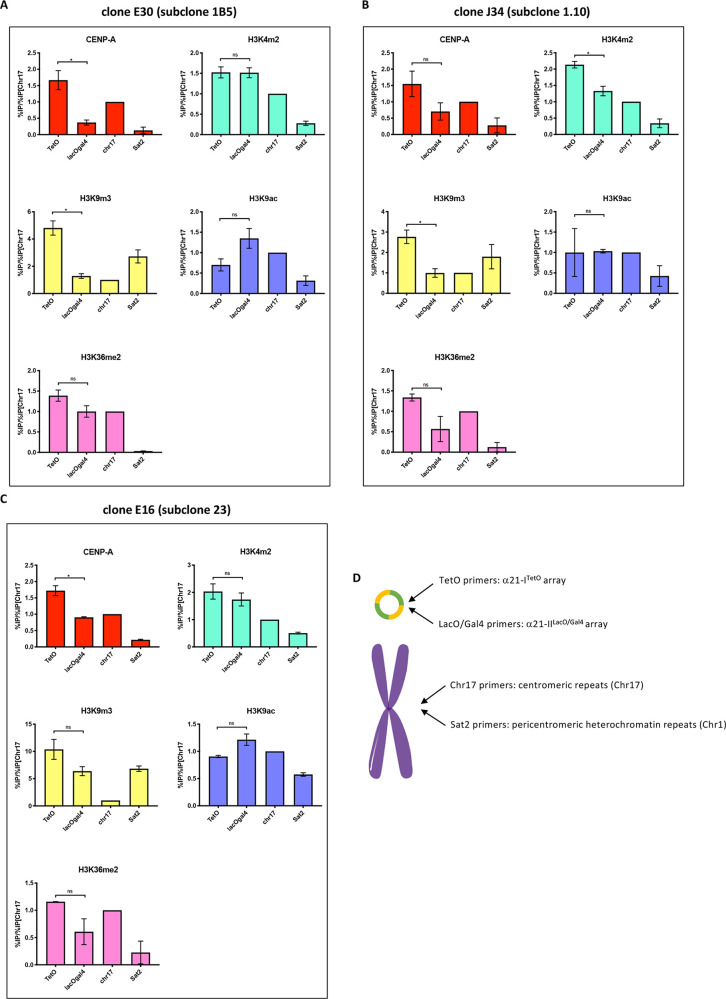
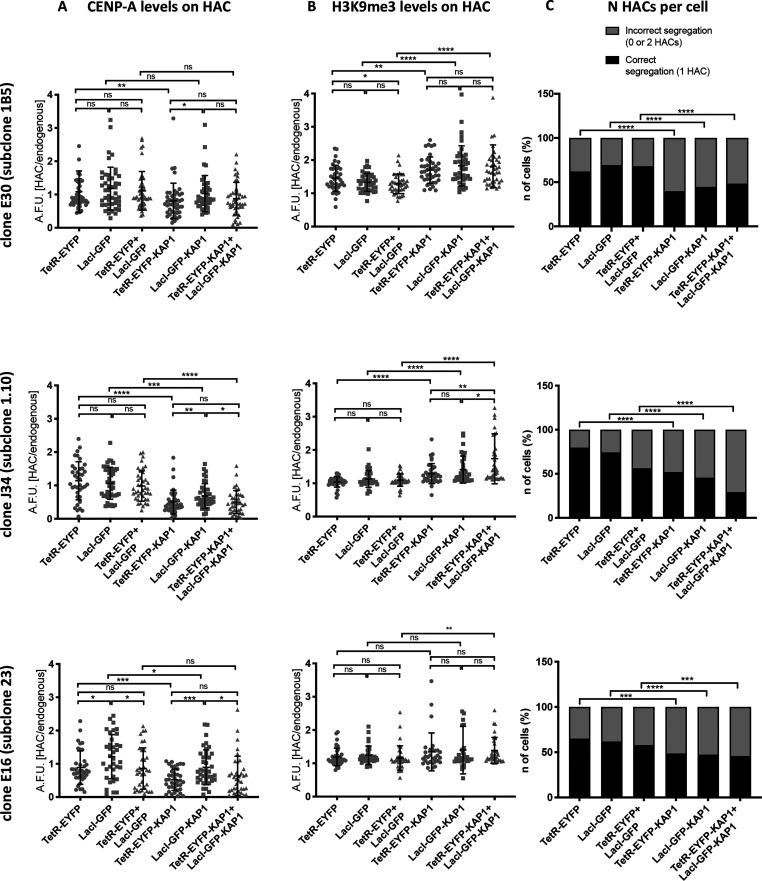
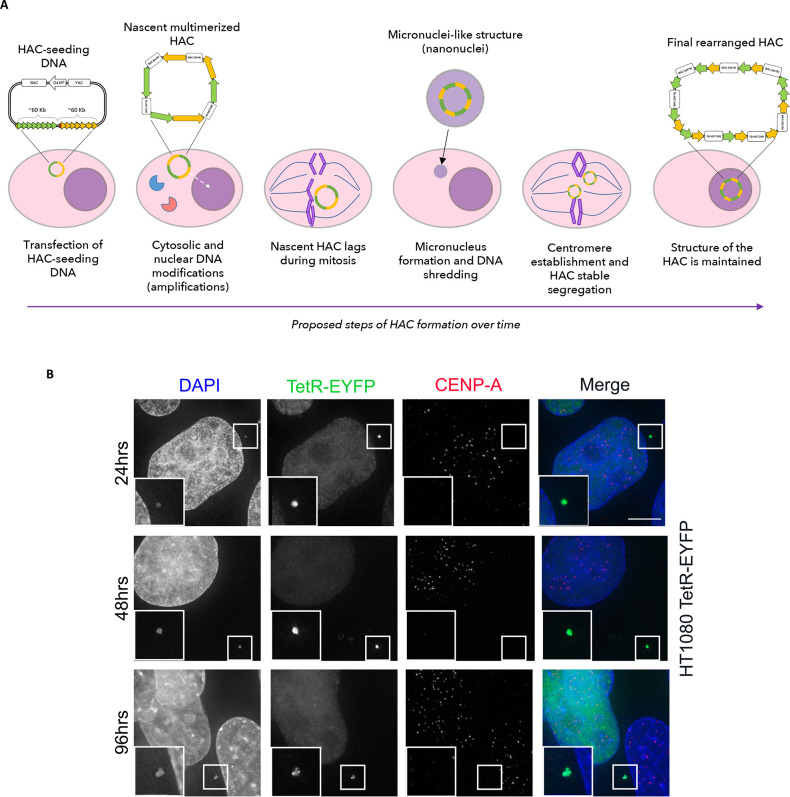
References
-
- Kouprina N.; Samoshkin A.; Erliandri I.; Nakano M.; Lee H. S.; Fu H.; Iida Y.; Aladjem M.; Oshimura M.; Masumoto H.; Earnshaw W. C.; Larionov V. (2012) Organization of Synthetic Alphoid DNA Array in Human Artificial Chromosome (HAC) with a Conditional Centromere. ACS Synth. Biol. 1, 590. 10.1021/sb3000436. - DOI - PMC - PubMed
-
- Pesenti E.; Kouprina N.; Liskovykh M.; Aurich-Costa J.; Larionov V.; Masumoto H.; Earnshaw W. C.; Molina O. (2018) Generation of a Synthetic Human Chromosome with Two Centromeric Domains for Advanced Epigenetic Engineering Studies. ACS Synth. Biol. 7, 1116. 10.1021/acssynbio.8b00018. - DOI - PMC - PubMed
-
- Lee N. C. O.; Kim J. H.; Petrov N. S.; Lee H. S.; Masumoto H.; Earnshaw W. C.; Larionov V.; Kouprina N. (2018) Method to Assemble Genomic DNA Fragments or Genes on Human Artificial Chromosome with Regulated Kinetochore Using a Multi-Integrase System. ACS Synth. Biol. 7, 63. 10.1021/acssynbio.7b00209. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
