A systems biology framework integrating GWAS and RNA-seq to shed light on the molecular basis of sperm quality in swine
- PMID: 33292187
- PMCID: PMC7724732
- DOI: 10.1186/s12711-020-00592-0
A systems biology framework integrating GWAS and RNA-seq to shed light on the molecular basis of sperm quality in swine
Abstract
Background: Genetic pressure in animal breeding is sparking the interest of breeders for selecting elite boars with higher sperm quality to optimize ejaculate doses and fertility rates. However, the molecular basis of sperm quality is not yet fully understood. Our aim was to identify candidate genes, pathways and DNA variants associated to sperm quality in swine by analysing 25 sperm-related phenotypes and integrating genome-wide association studies (GWAS) and RNA-seq under a systems biology framework.
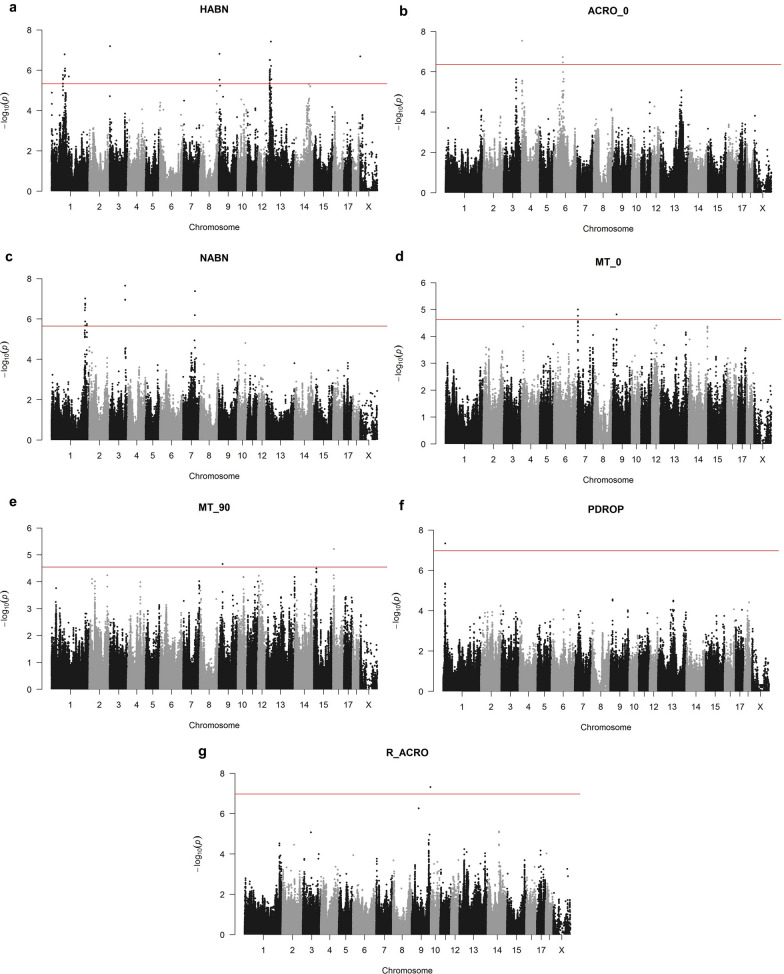
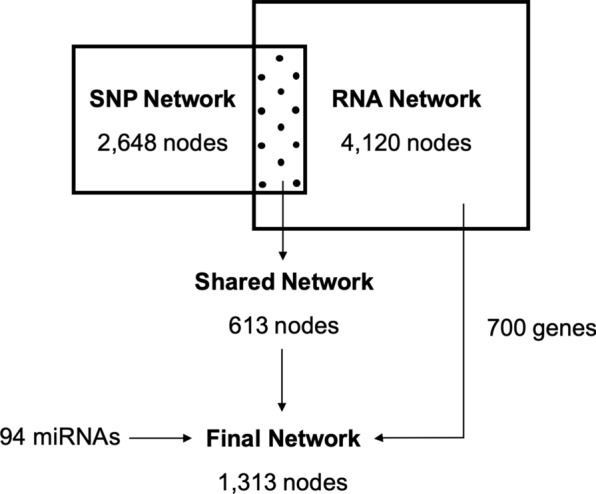
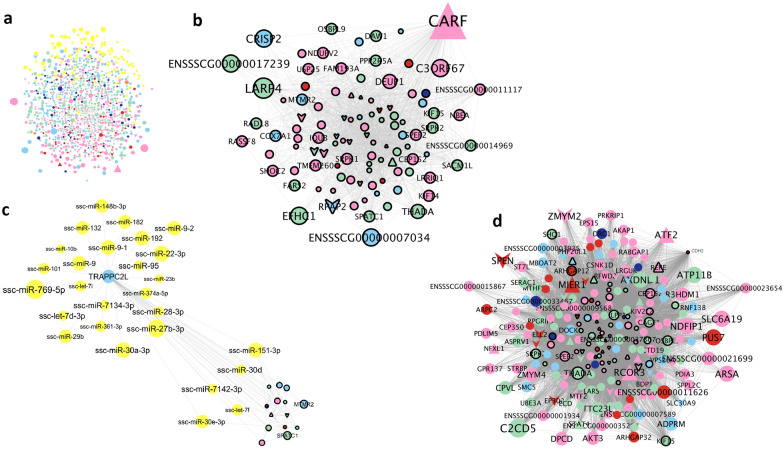
Results: By GWAS, we identified 12 quantitative trait loci (QTL) associated to the percentage of head and neck abnormalities, abnormal acrosomes and motile spermatozoa. Candidate genes included CHD2, KATNAL2, SLC14A2 and ABCA1. By RNA-seq, we identified a wide repertoire of mRNAs (e.g. PRM1, OAZ3, DNAJB8, TPPP2 and TNP1) and miRNAs (e.g. ssc-miR-30d, ssc-miR-34c, ssc-miR-30c-5p, ssc-miR-191, members of the let-7 family and ssc-miR-425-5p) with functions related to sperm biology. We detected 6128 significant correlations (P-value ≤ 0.05) between sperm traits and mRNA abundances. By expression (e)GWAS, we identified three trans-expression QTL involving the genes IQCJ, ACTR2 and HARS. Using the GWAS and RNA-seq data, we built a gene interaction network. We considered that the genes and interactions that were present in both the GWAS and RNA-seq networks had a higher probability of being actually involved in sperm quality and used them to build a robust gene interaction network. In addition, in the final network we included genes with RNA abundances correlated with more than four semen traits and miRNAs interacting with the genes on the network. The final network was enriched for genes involved in gamete generation and development, meiotic cell cycle, DNA repair or embryo implantation. Finally, we designed a panel of 73 SNPs based on the GWAS, eGWAS and final network data, that explains between 5% (for sperm cell concentration) and 36% (for percentage of neck abnormalities) of the phenotypic variance of the sperm traits.
Conclusions: By applying a systems biology approach, we identified genes that potentially affect sperm quality and constructed a SNP panel that explains a substantial part of the phenotypic variance for semen quality in our study and that should be tested in other swine populations to evaluate its relevance for the pig breeding sector.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures