

CXCR7: a β-arrestin-biased receptor that potentiates cell migration and recruits β-arrestin2 exclusively through Gβγ subunits and GRK2
- PMID: 33292475
- PMCID: PMC7686738
- DOI: 10.1186/s13578-020-00497-x
CXCR7: a β-arrestin-biased receptor that potentiates cell migration and recruits β-arrestin2 exclusively through Gβγ subunits and GRK2
Abstract
Background: Some chemokine receptors referred to as atypical chemokine receptors (ACKRs) are thought to non-signaling decoys because of their inability to activate typical G-protein signaling pathways. CXCR7, also known as ACKR3, binds to only two chemokines, SDF-1α and I-TAC, and recruits β-arrestins. SDF-1α also binds to its own conventional receptor, CXCR4, involving in homeostatic modulation such as development and immune surveillance as well as pathological conditions such as inflammation, ischemia, and cancers. Recently, CXCR7 is suggested as a key therapeutic target together with CXCR4 in such conditions. However, the molecular mechanisms underlying cellular responses and functional relation with CXCR7 and CXCR4 have not been elucidated, despite massive studies. Therefore, we aimed to reveal the molecular networks of CXCR7 and CXCR4 and compare their effects on cell migration.
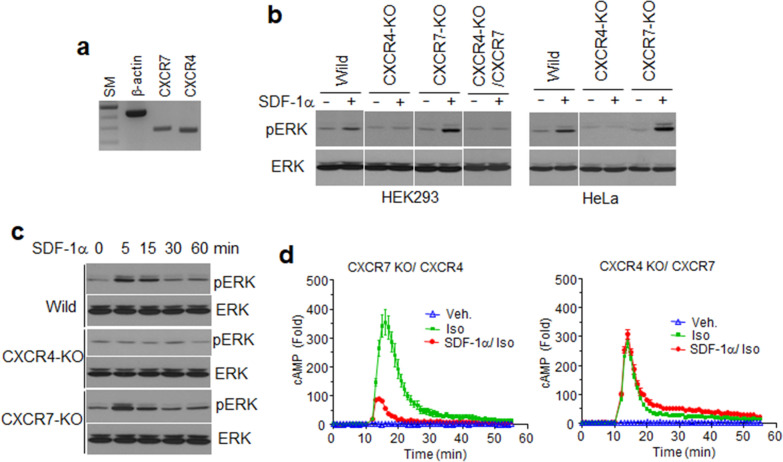
Methods: Base on structural complementation assay using NanoBiT technology, we characterized the distinct mechanisms underlying β-arrestin2 recruitment by both CXCR4 and CXCR7. Crosslinking and immunoprecipitation were conducted to analyze complex formation of the receptors. Gene deletion using CRISPR and reconstitution of the receptors were applied to analysis of ligand-dependent ERK phosphorylation and cell migration. All experiments were performed in triplicate and repeated more than three times. Unpaired Student's t-tests or ANOVA using PRISM5 software were employed for statistical analyses.
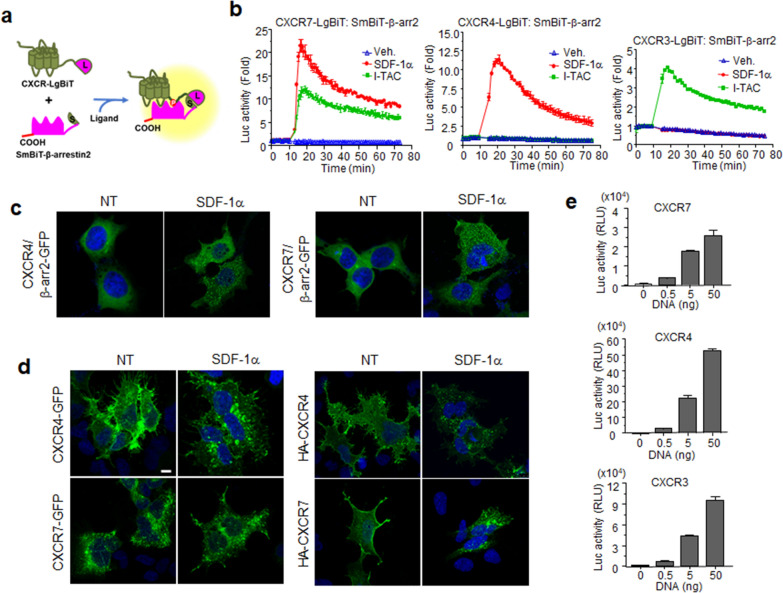
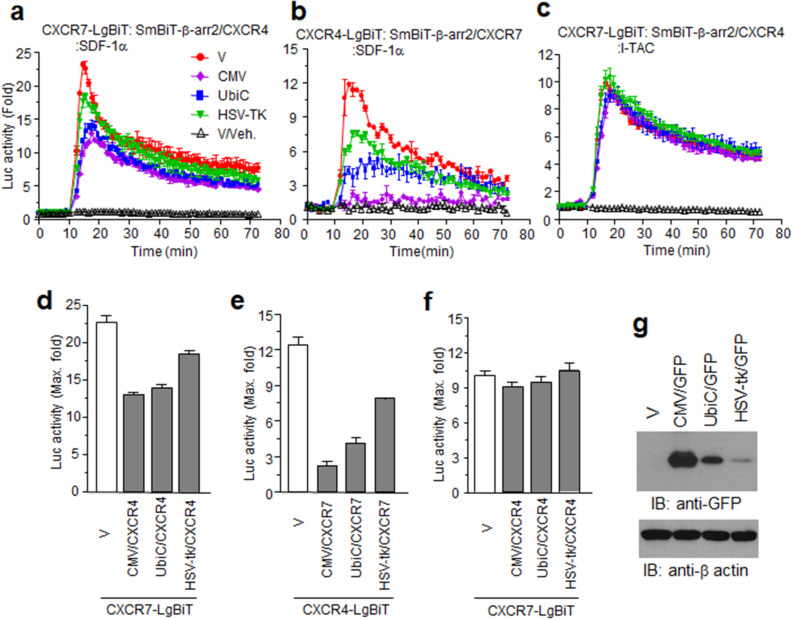
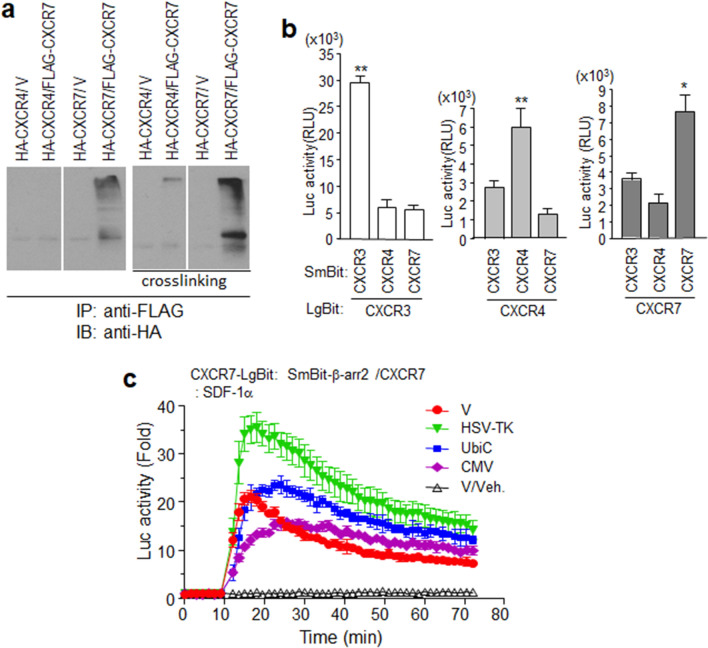
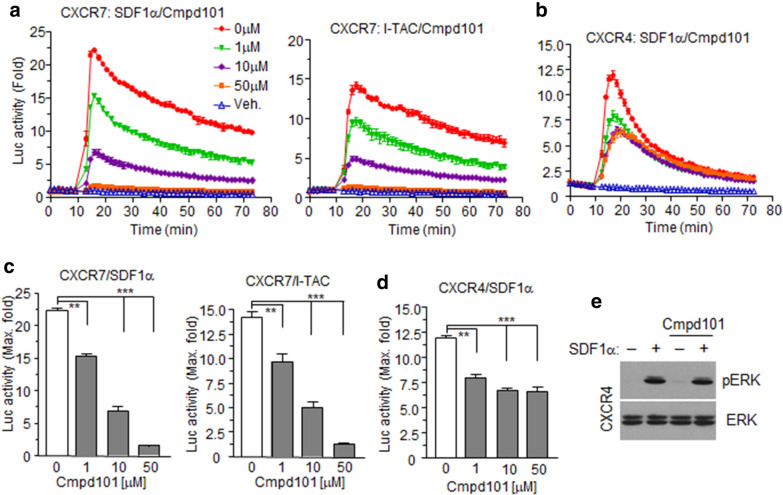
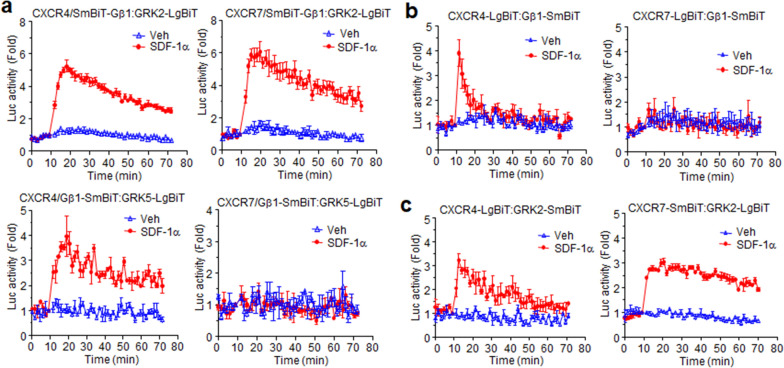
Results: Ligand binding to CXCR7 does not result in activation of typical signaling pathways via Gα subunits but activation of GRK2 via βγ subunits and receptor phosphorylation with subsequent β-arrestin2 recruitment. In contrast, CXCR4 induced Gαi activation and recruited β-arrestin2 through C-terminal phosphorylation by both GRK2 and GRK5. SDF-1α-stimulated ERK phosphorylation was facilitated by CXCR4, but not CXCR7. Heterodimerization of CXCR4 and CXCR7 was not confirmed in this study, while homodimerization of them was verified by crosslinking experiment and NanoBiT assay. Regarding chemotaxis, SDF-1α-stimulated cell migration was mediated by both CXCR4 and CXCR7.
Conclusion: This study demonstrates that SDF-1α-stimulated CXCR7 mediates β-arrestin2 recruitment via different molecular networking from that of CXCR4. CXCR7 may be neither a simple scavenger nor auxiliary receptor but plays an essential role in cell migration through cooperation with CXCR4.
Keywords: Biased GPCR; CXCR4; CXCR7; Chemotaxis; SDF-1α; Structural complementation assay.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
-
- Bachelerie F, Ben-Baruch A, Burkhardt AM, Combadiere C, Farber JM, Graham GJ, Horuk R, Sparre-Ulrich AH, Locati M, Luster AD, Mantovani A, Matsushima K, Murphy PM, Nibbs R, Nomiyama H, Power CA, Proudfoot AE, Rosenkilde MM, Rot A, Sozzani S, Thelen M, Yoshie O, Zlotnik A. International Union of Basic and Clinical Pharmacology. [corrected]. LXXXIX. Update on the extended family of chemokine receptors and introducing a new nomenclature for atypical chemokine receptors. Pharmacol Rev. 2014;66(1):1–79. doi: 10.1124/pr.113.007724. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
