

Dynamics of RNA polymerase II and elongation factor Spt4/5 recruitment during activator-dependent transcription
- PMID: 33293419
- PMCID: PMC7768755
- DOI: 10.1073/pnas.2011224117
Dynamics of RNA polymerase II and elongation factor Spt4/5 recruitment during activator-dependent transcription
Abstract
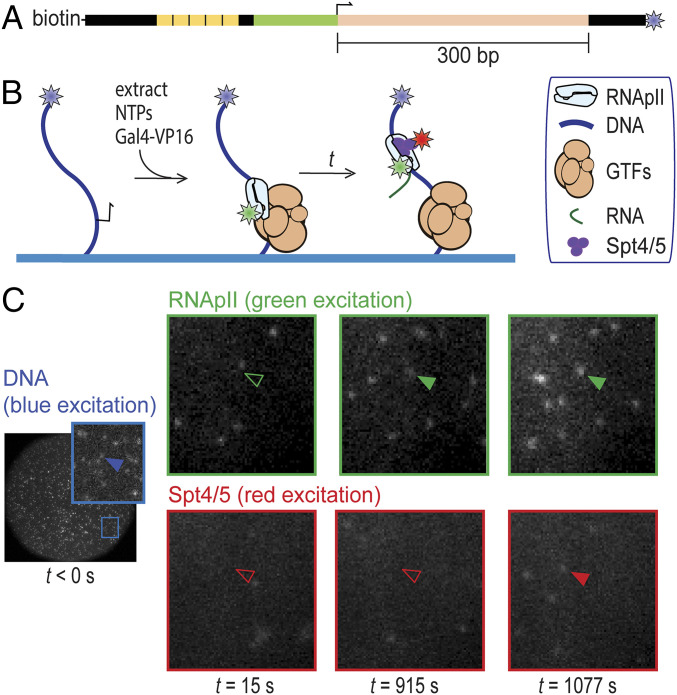
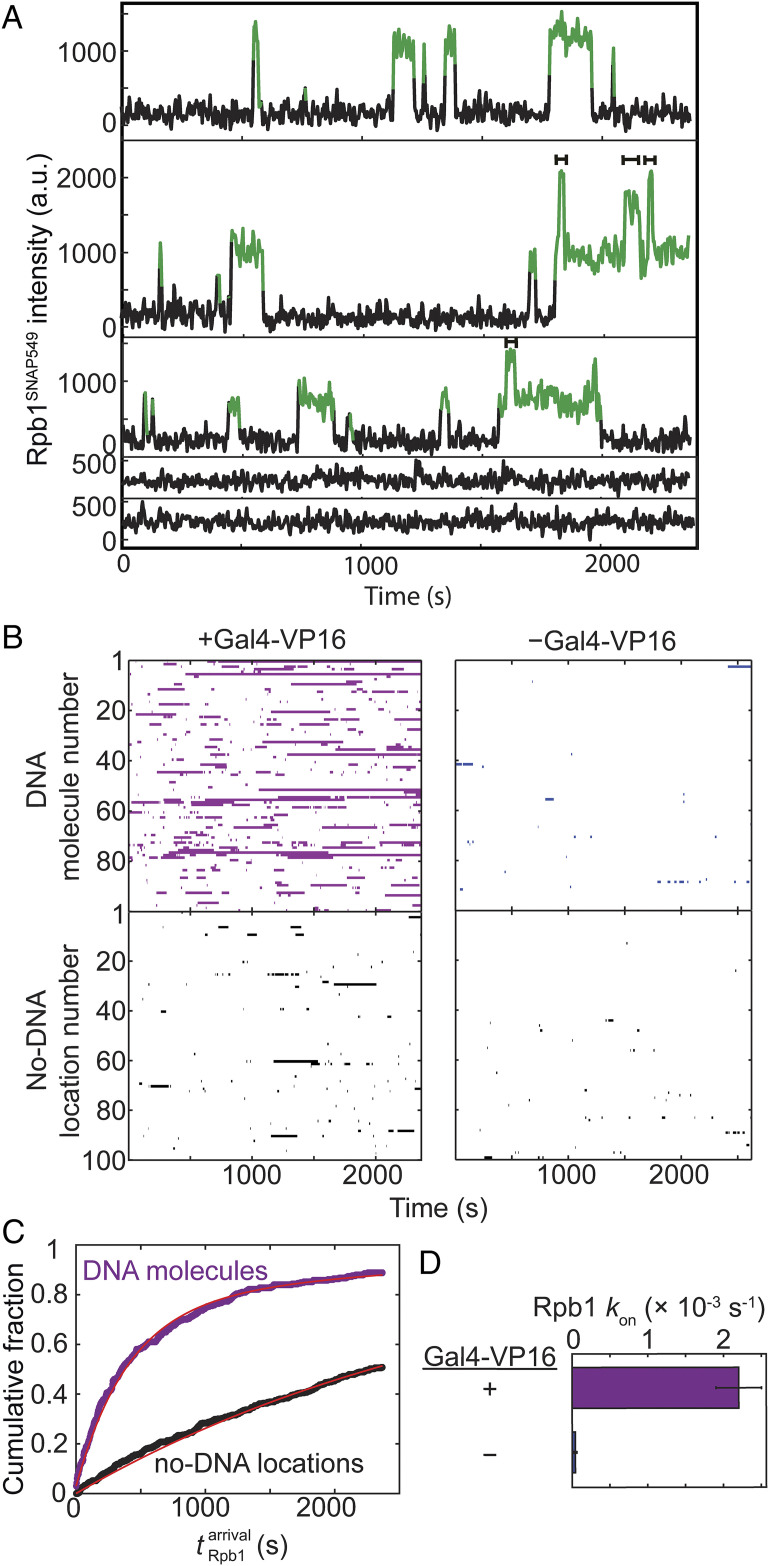
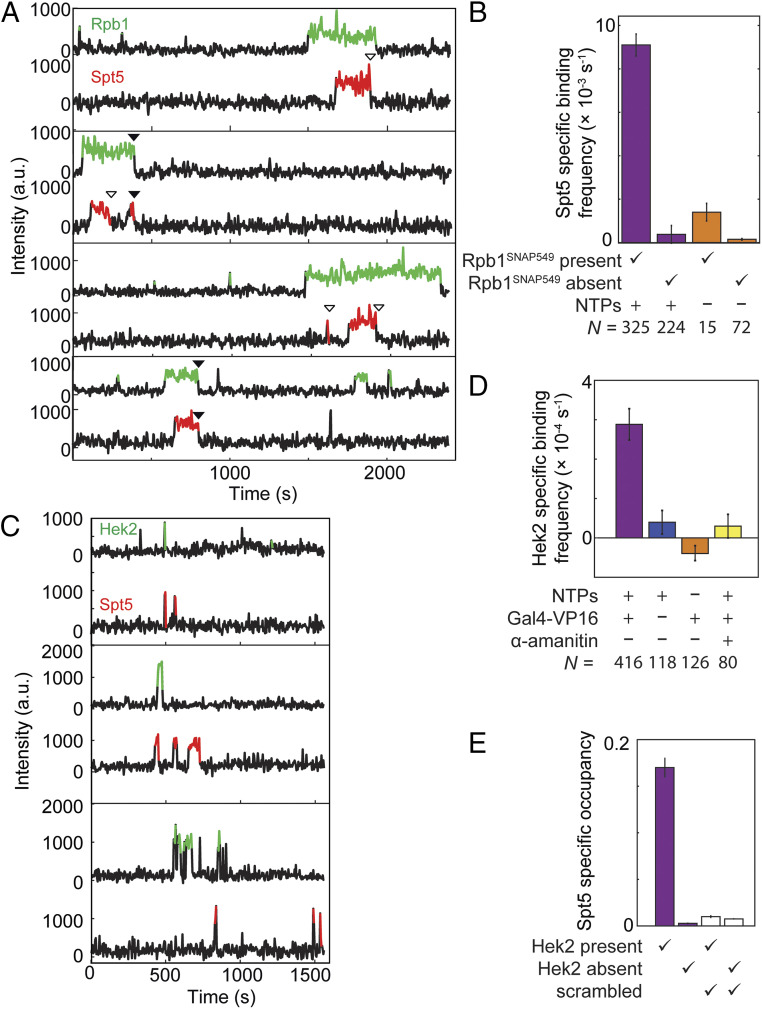
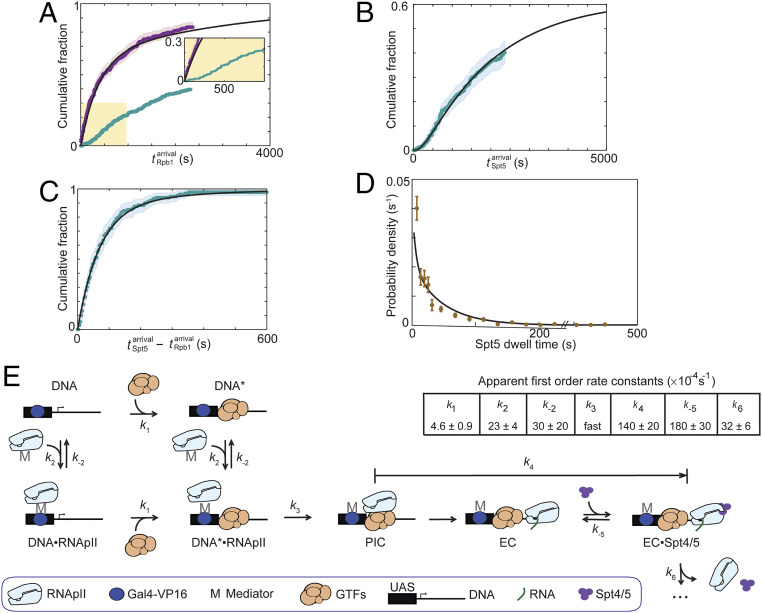
In eukaryotes, RNA polymerase II (RNApII) transcribes messenger RNA from template DNA. Decades of experiments have identified the proteins needed for transcription activation, initiation complex assembly, and productive elongation. However, the dynamics of recruitment of these proteins to transcription complexes, and of the transitions between these steps, are poorly understood. We used multiwavelength single-molecule fluorescence microscopy to directly image and quantitate these dynamics in a budding yeast nuclear extract that reconstitutes activator-dependent transcription in vitro. A strong activator (Gal4-VP16) greatly stimulated reversible binding of individual RNApII molecules to template DNA. Binding of labeled elongation factor Spt4/5 to DNA typically followed RNApII binding, was NTP dependent, and was correlated with association of mRNA binding protein Hek2, demonstrating specificity of Spt4/5 binding to elongation complexes. Quantitative kinetic modeling shows that only a fraction of RNApII binding events are productive and implies a rate-limiting step, probably associated with recruitment of general transcription factors, needed to assemble a transcription-competent preinitiation complex at the promoter. Spt4/5 association with transcription complexes was slowly reversible, with DNA-bound RNApII molecules sometimes binding and releasing Spt4/5 multiple times. The average Spt4/5 residence time was of similar magnitude to the time required to transcribe an average length yeast gene. These dynamics suggest that a single Spt4/5 molecule remains associated during a typical transcription event, yet can dissociate from RNApII to allow disassembly of abnormally long-lived (i.e., stalled) elongation complexes.
Keywords: CoSMoS; Gal4-VP16; Saccharomyces cerevisiae.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Thomas M. C., Chiang C. M., The general transcription machinery and general cofactors. Crit. Rev. Biochem. Mol. Biol. 41, 105–178 (2006). - PubMed
-
- Mayer A., et al. , Uniform transitions of the general RNA polymerase II transcription complex. Nat. Struct. Mol. Biol. 17, 1272–1278 (2010). - PubMed
-
- Vos S. M., et al. , Structure of activated transcription complex Pol II-DSIF-PAF-SPT6. Nature 560, 607–612 (2018). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
