

Human T-bet Governs Innate and Innate-like Adaptive IFN-γ Immunity against Mycobacteria
- PMID: 33296702
- PMCID: PMC7770098
- DOI: 10.1016/j.cell.2020.10.046
Human T-bet Governs Innate and Innate-like Adaptive IFN-γ Immunity against Mycobacteria
Abstract
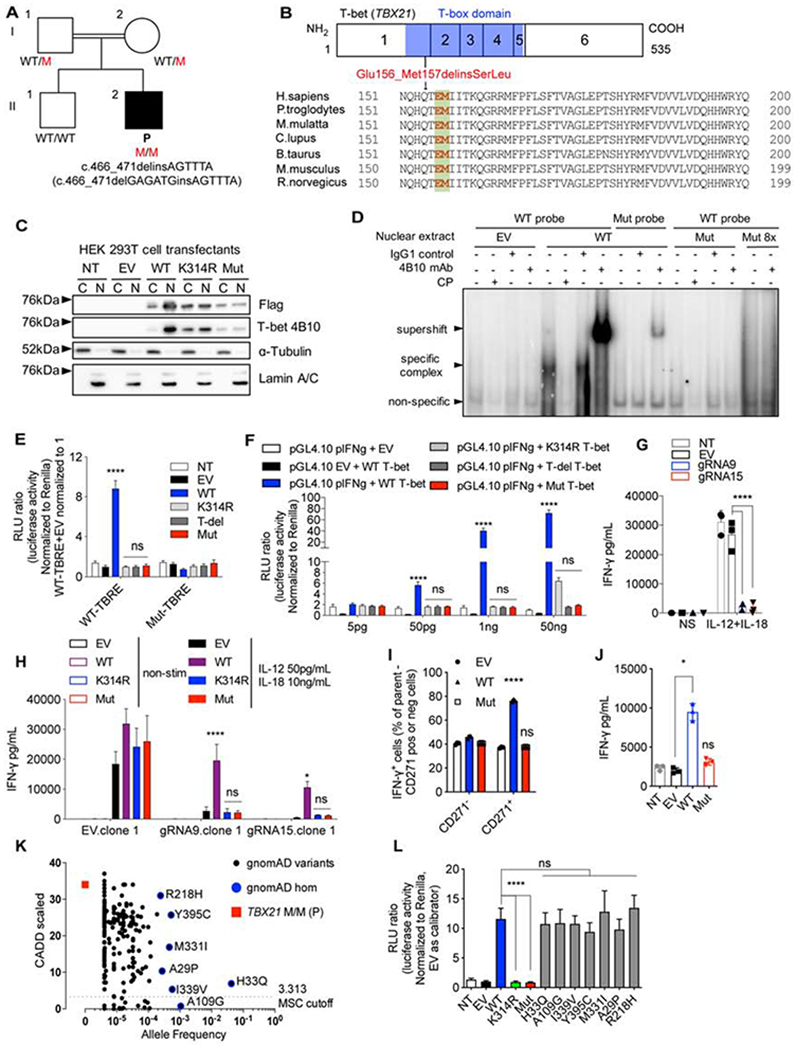
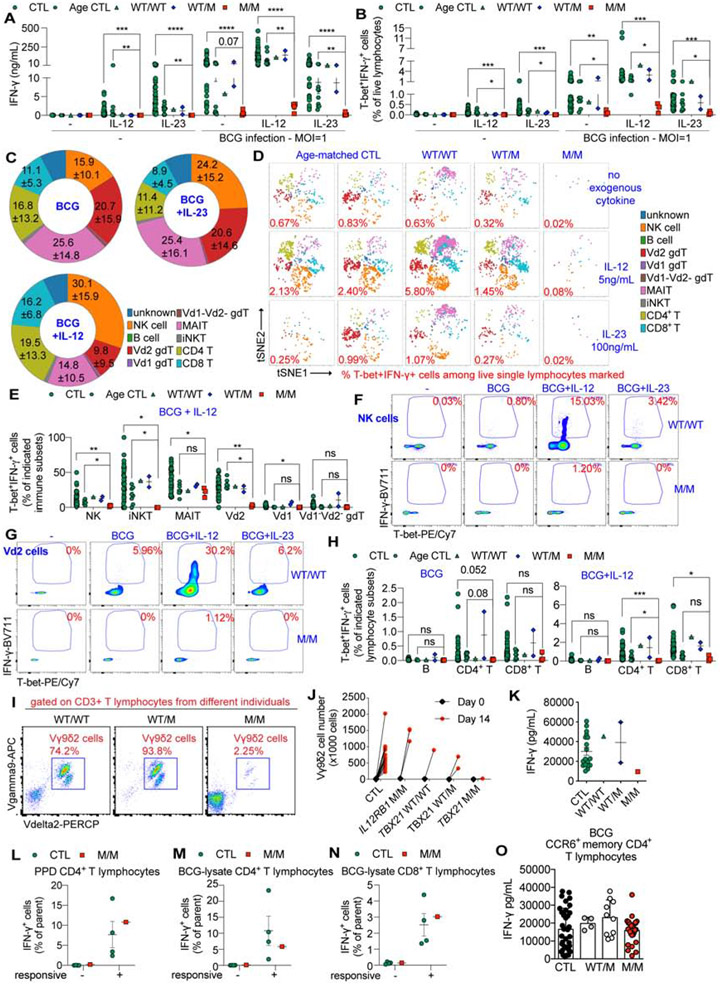
Inborn errors of human interferon gamma (IFN-γ) immunity underlie mycobacterial disease. We report a patient with mycobacterial disease due to inherited deficiency of the transcription factor T-bet. The patient has extremely low counts of circulating Mycobacterium-reactive natural killer (NK), invariant NKT (iNKT), mucosal-associated invariant T (MAIT), and Vδ2+ γδ T lymphocytes, and of Mycobacterium-non reactive classic TH1 lymphocytes, with the residual populations of these cells also producing abnormally small amounts of IFN-γ. Other lymphocyte subsets develop normally but produce low levels of IFN-γ, with the exception of CD8+ αβ T and non-classic CD4+ αβ TH1∗ lymphocytes, which produce IFN-γ normally in response to mycobacterial antigens. Human T-bet deficiency thus underlies mycobacterial disease by preventing the development of innate (NK) and innate-like adaptive lymphocytes (iNKT, MAIT, and Vδ2+ γδ T cells) and IFN-γ production by them, with mycobacterium-specific, IFN-γ-producing, purely adaptive CD8+ αβ T, and CD4+ αβ TH1∗ cells unable to compensate for this deficit.
Keywords: IFN-γ; Mendelian susceptibility to mycobacterial disease; T-bet; immunodeficiency; inborn errors of immunity; innate lymphocyte; innate-like adaptive lymphocyte; mycobacterium.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests L.H.G. serves on the Board of Directors of GlaxoSmithKline Pharmaceutical Company and the Analog Device Corporation and formerly served on the Boards of Bristol Myers Squibb Pharmaceutical Company and the Waters Corporation. She is also on the Scientific Advisory Boards of Abpro, Kaleido, and Repare biotechnology companies. J.-L.C. serves on the Scientific Advisory Boards of ADMA Biologics Inc., Celgene, and Kymera Therapeutics, Inc. He also consults for Elixiron Immunotherapeutics. Other authors declare no competing interests.
Figures




References
-
- Abecasis GR, Cherny SS, Cookson WO, and Cardon LR (2002). Merlin—rapid analysis of dense genetic maps using sparse gene flow trees. Nat. Genet 30, 97–101. - PubMed
-
- Acosta-Rodriguez EV, Rivino L, Geginat J, Jarrossay D, Gattorno M, Lanzavecchia A, Sallusto F, and Napolitani G (2007). Surface phenotype and antigenic specificity of human interleukin 17-producing T helper memory cells. Nat. Immunol 8, 639–646. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
