

The involvement of neuroimmune cells in adipose innervation
- PMID: 33297933
- PMCID: PMC7727151
- DOI: 10.1186/s10020-020-00254-3
The involvement of neuroimmune cells in adipose innervation
Abstract
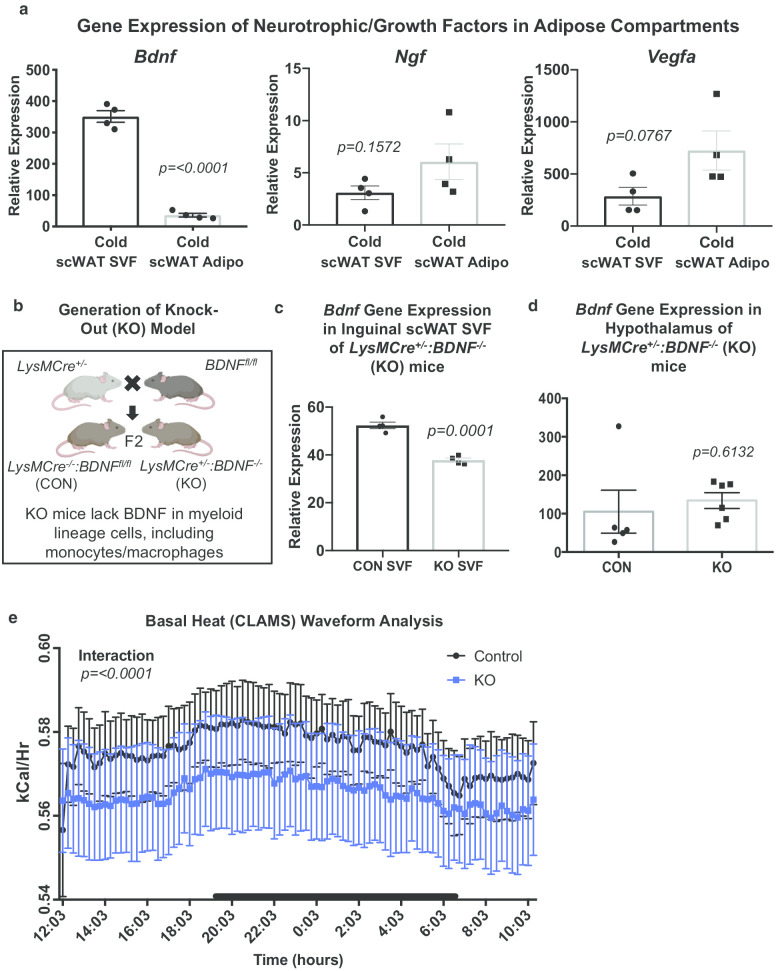
Background: Innervation of adipose tissue is essential for the proper function of this critical metabolic organ. Numerous surgical and chemical denervation studies have demonstrated how maintenance of brain-adipose communication through both sympathetic efferent and sensory afferent nerves helps regulate adipocyte size, cell number, lipolysis, and 'browning' of white adipose tissue. Neurotrophic factors are growth factors that promote neuron survival, regeneration, and plasticity, including neurite outgrowth and synapse formation. Peripheral immune cells have been shown to be a source of neurotrophic factors in humans and mice. Although a number of immune cells reside in the adipose stromal vascular fraction (SVF), it has remained unclear what roles they play in adipose innervation. We previously demonstrated that adipose SVF secretes brain derived neurotrophic factor (BDNF).
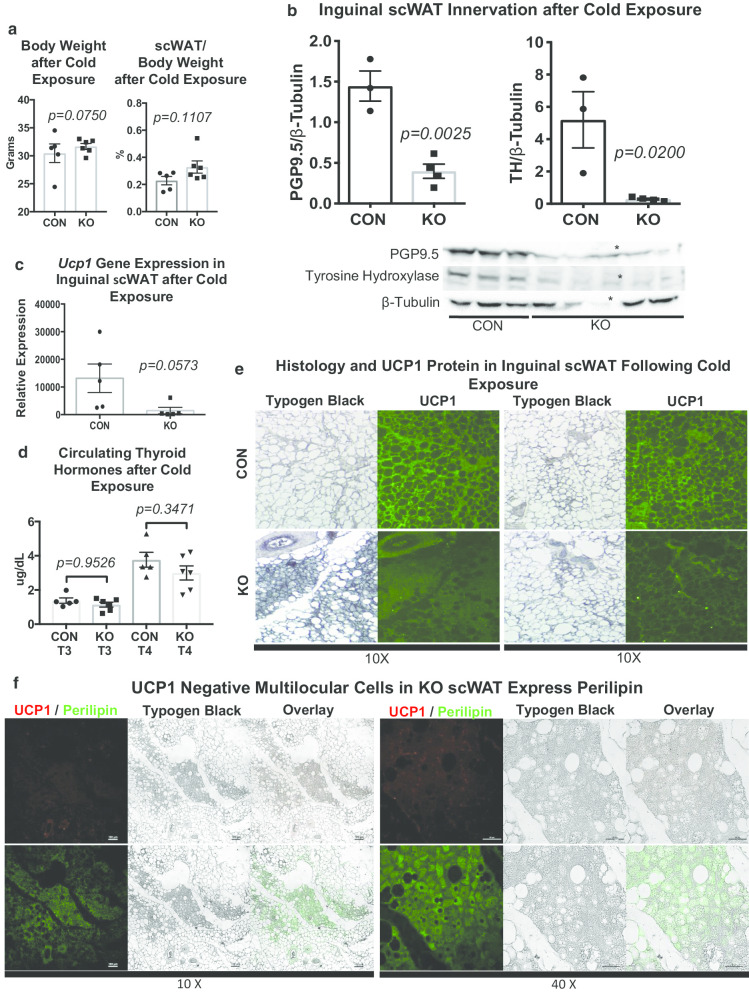
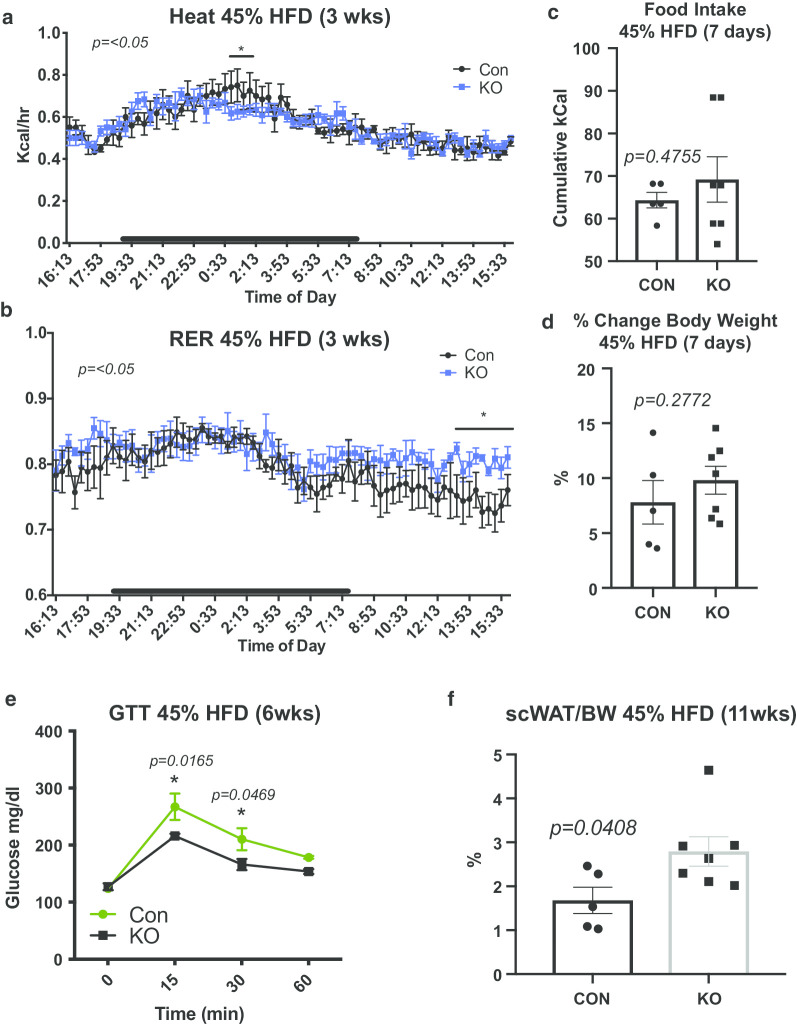
Methods: We now show that deletion of this neurotrophic factor from the myeloid lineage of immune cells led to a 'genetic denervation' of inguinal subcutaneous white adipose tissue (scWAT), thereby causing decreased energy expenditure, increased adipose mass, and a blunted UCP1 response to cold stimulation.
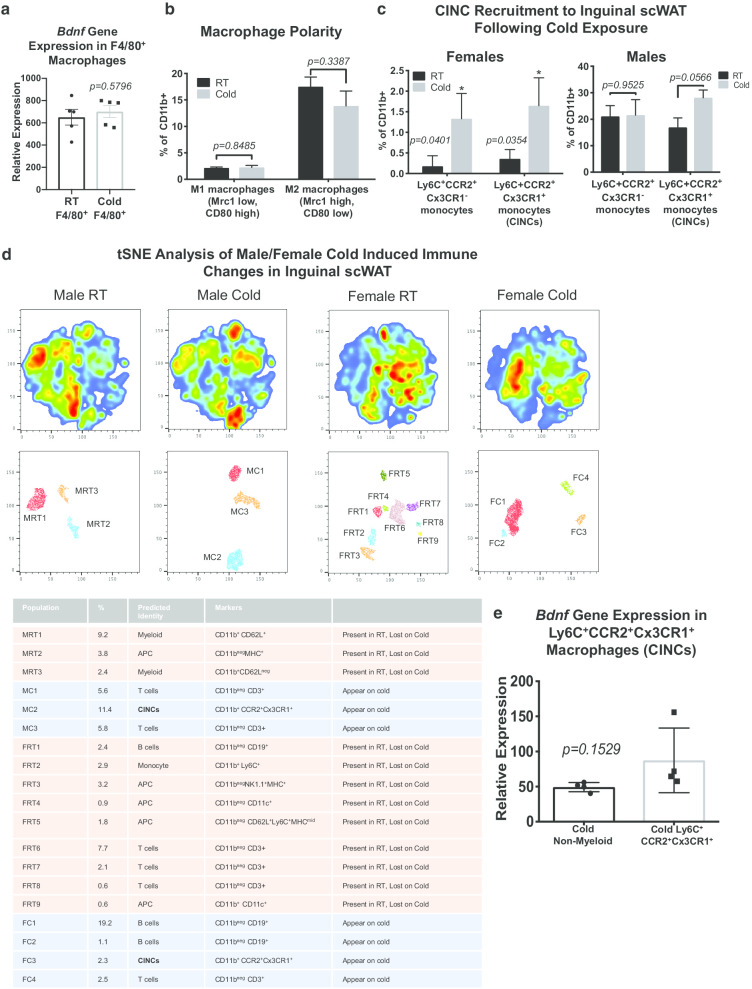
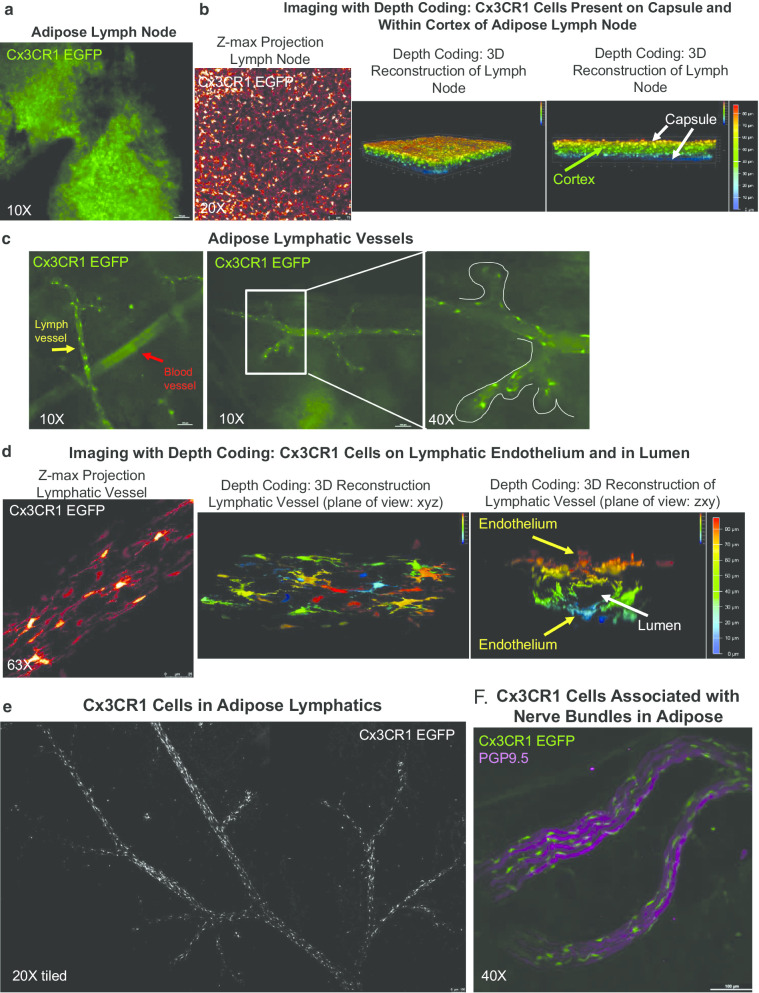
Results: We and others have previously shown that noradrenergic stimulation via cold exposure increases adipose innervation in the inguinal depot. Here we have identified a subset of myeloid cells that home to scWAT upon cold exposure and are Ly6C+ CCR2+ Cx3CR1+ monocytes/macrophages that express noradrenergic receptors and BDNF. This subset of myeloid lineage cells also clearly interacted with peripheral nerves in the scWAT and were therefore considered neuroimmune cells.
Conclusions: We propose that these myeloid lineage, cold induced neuroimmune cells (CINCs) are key players in maintaining adipose innervation as well as promoting adipose nerve remodeling under noradrenergic stimulation, such as cold exposure.
Keywords: BDNF; Browning; CINCs; Cold-induced neuroimmune cells; Energy expenditure; Innervation; Monocyte/macrophage; White adipose tissue (WAT).
Conflict of interest statement
The authors do not have any competing interests to disclose.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
