

Preexisting immunity shapes distinct antibody landscapes after influenza virus infection and vaccination in humans
- PMID: 33298562
- PMCID: PMC8115023
- DOI: 10.1126/scitranslmed.abd3601
Preexisting immunity shapes distinct antibody landscapes after influenza virus infection and vaccination in humans
Abstract
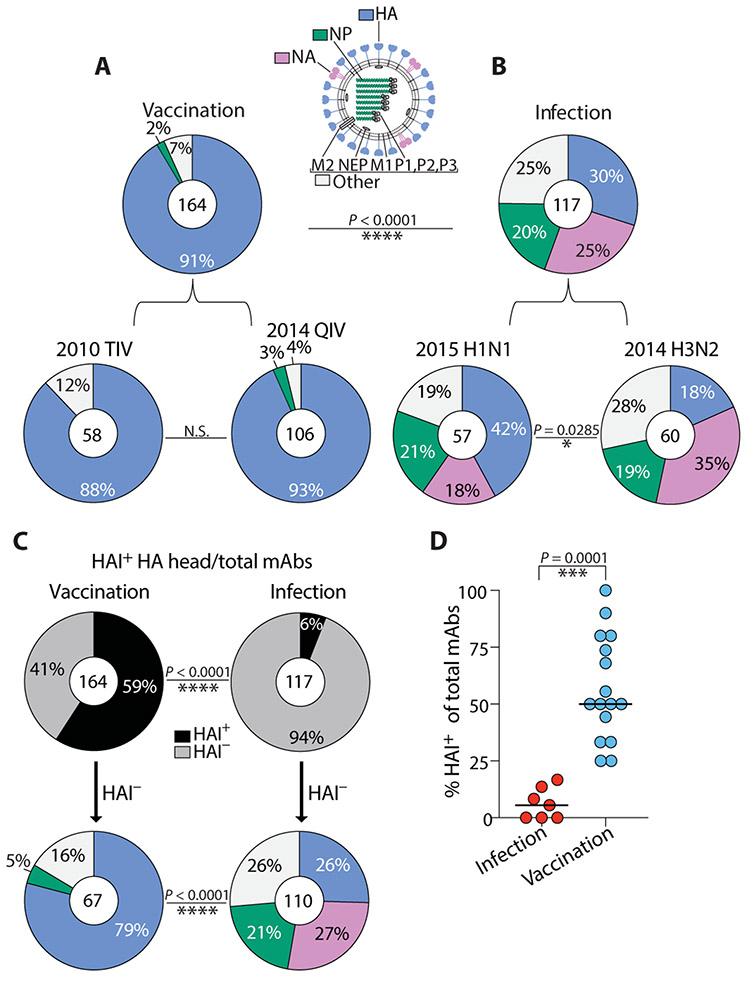
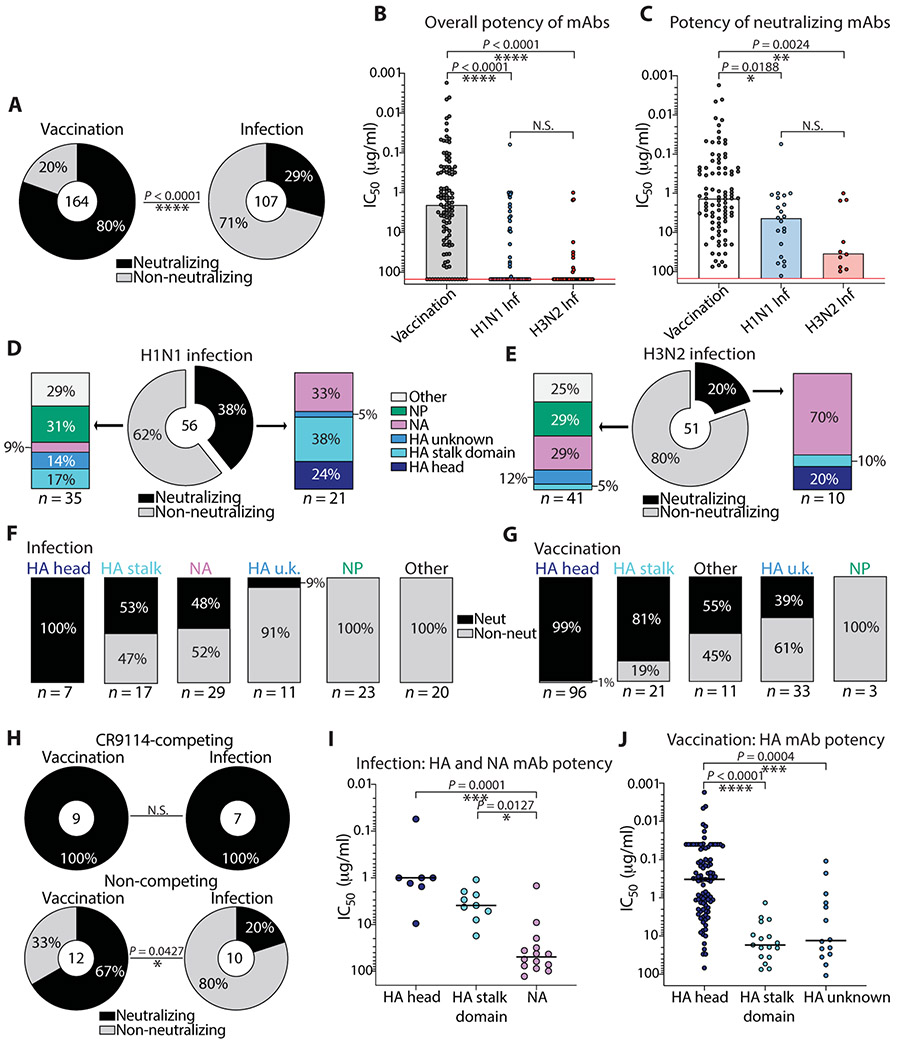
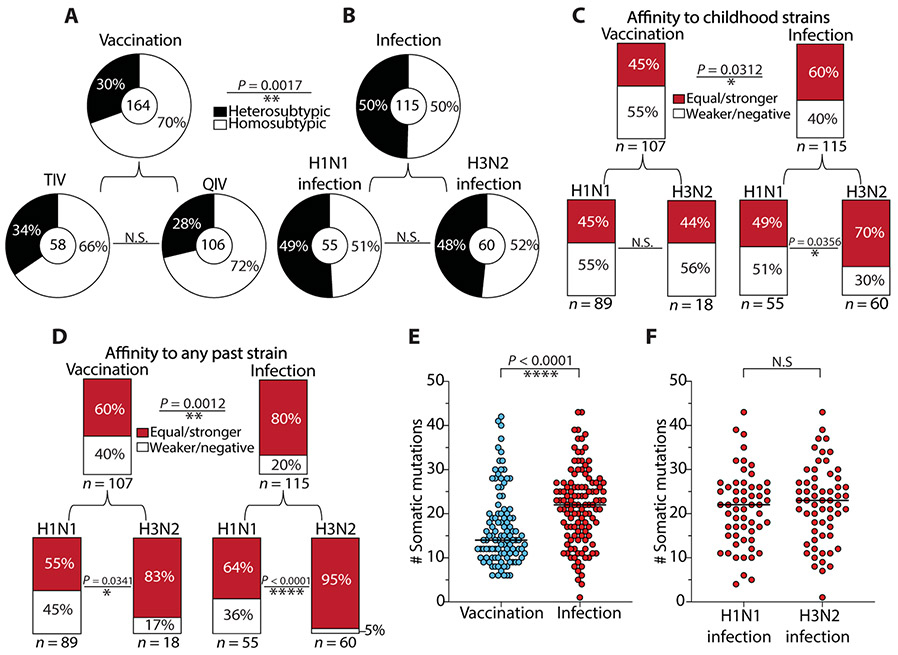
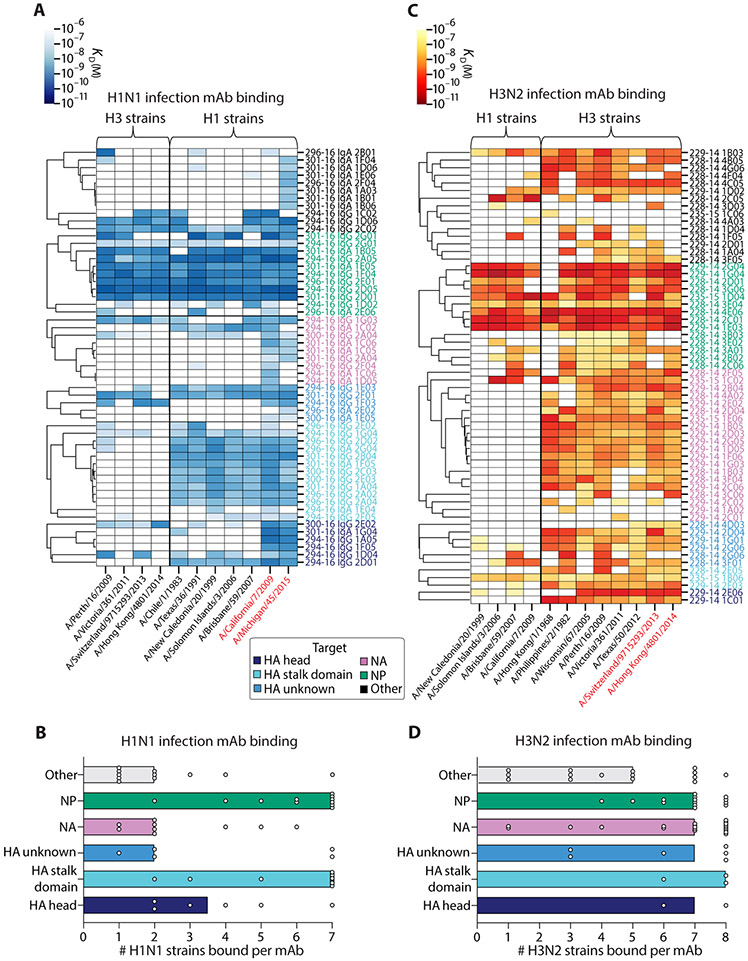
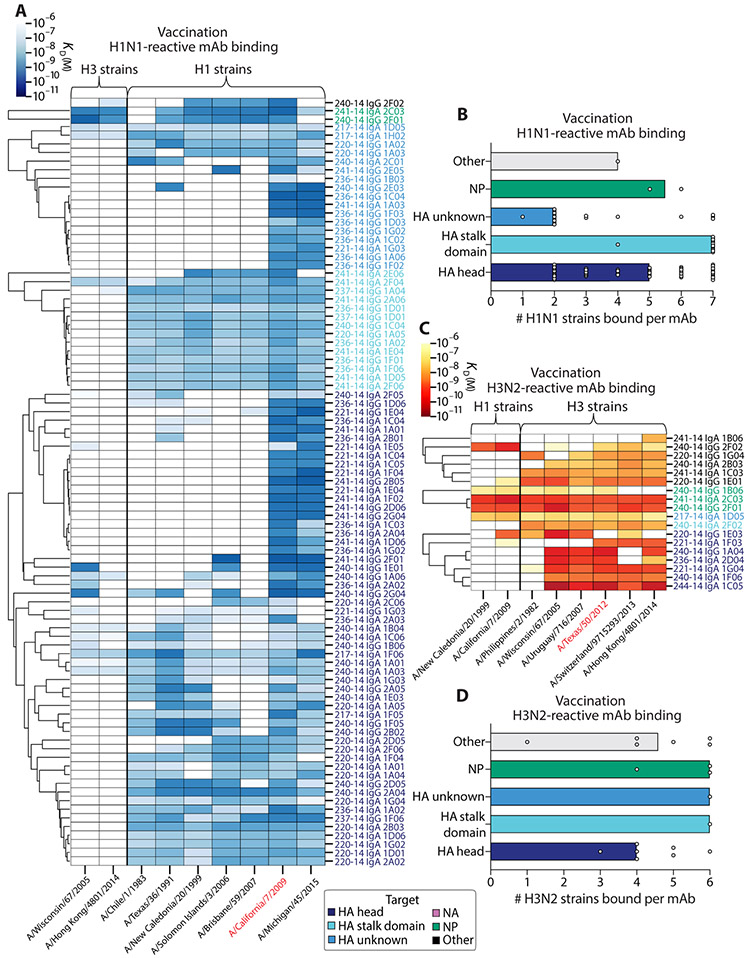
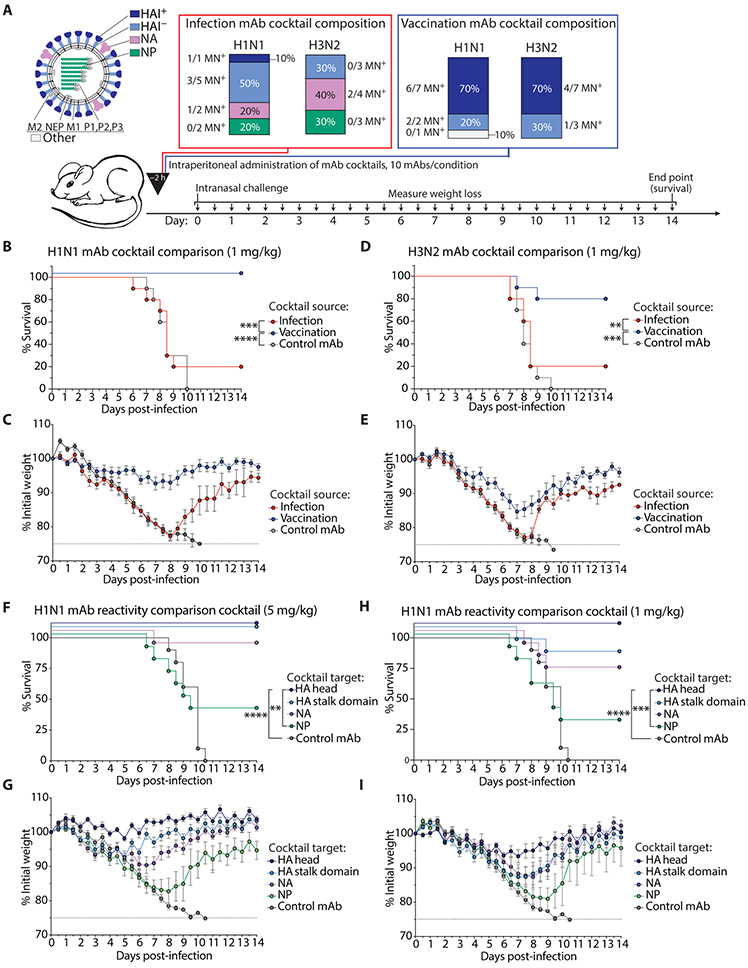
Humans are repeatedly exposed to variants of influenza virus throughout their lifetime. As a result, preexisting influenza-specific memory B cells can dominate the response after infection or vaccination. Memory B cells recalled by adulthood exposure are largely reactive to conserved viral epitopes present in childhood strains, posing unclear consequences on the ability of B cells to adapt to and neutralize newly emerged strains. We sought to investigate the impact of preexisting immunity on generation of protective antibody responses to conserved viral epitopes upon influenza virus infection and vaccination in humans. We accomplished this by characterizing monoclonal antibodies (mAbs) from plasmablasts, which are predominantly derived from preexisting memory B cells. We found that, whereas some influenza infection-induced mAbs bound conserved and neutralizing epitopes on the hemagglutinin (HA) stalk domain or neuraminidase, most of the mAbs elicited by infection targeted non-neutralizing epitopes on nucleoprotein and other unknown antigens. Furthermore, most infection-induced mAbs had equal or stronger affinity to childhood strains, indicating recall of memory B cells from childhood exposures. Vaccination-induced mAbs were similarly induced from past exposures and exhibited substantial breadth of viral binding, although, in contrast to infection-induced mAbs, they targeted neutralizing HA head epitopes. Last, cocktails of infection-induced mAbs displayed reduced protective ability in mice compared to vaccination-induced mAbs. These findings reveal that both preexisting immunity and exposure type shape protective antibody responses to conserved influenza virus epitopes in humans. Natural infection largely recalls cross-reactive memory B cells against non-neutralizing epitopes, whereas vaccination harnesses preexisting immunity to target protective HA epitopes.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
References
-
- Iuliano AD, Roguski KM, Chang HH, Muscatello DJ, Palekar R, Tempia S, Cohen C, Gran JM, Schanzer D, Cowling BJ, Wu P, Kyncl J, Ang LW, Park M, Redlberger-Fritz M, Yu H, Espenhain L, Krishnan A, Emukule G, van Asten L, Pereira da Silva S, Aungkulanon S, Buchholz U, Widdowson M-A, Bresee JS; Global Seasonal Influenza-associated Mortality Collaborator Network, Estimates of global seasonal influenza-associated respiratory mortality: A modelling study. Lancet 391, 1285–1300 (2018). - PMC - PubMed
-
- Dawood FS, Chung JR, Kim SS, Zimmerman RK, Nowalk MP, Jackson ML, Jackson LA, Monto AS, Martin ET, Belongia EA, McLean HQ, Gaglani M, Dunnigan K, Foust A, Sessions W, DaSilva J, Le S, Stark T, Kondor RJ, Barnes JR, Wentworth DE, Brammer L, Fry AM, Patel MM, Flannery B, Interim estimates of 2019-20 seasonal influenza vaccine effectiveness—United States, February 2020. MMWR Morb. Mortal. Wkly Rep 69, 177–182 (2020). - PMC - PubMed
-
- Webster RG, Laver WG, Air GM, Schild GC, Molecular mechanisms of variation in influenza viruses. Nature 296, 115–121 (1982). - PubMed
-
- Krammer F, Palese P, Advances in the development of influenza virus vaccines. Nat. Rev. Drug Discov 14, 167–182 (2015). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
