

High-speed atomic force microscopy highlights new molecular mechanism of daptomycin action
- PMID: 33298927
- PMCID: PMC7725780
- DOI: 10.1038/s41467-020-19710-z
High-speed atomic force microscopy highlights new molecular mechanism of daptomycin action
Abstract
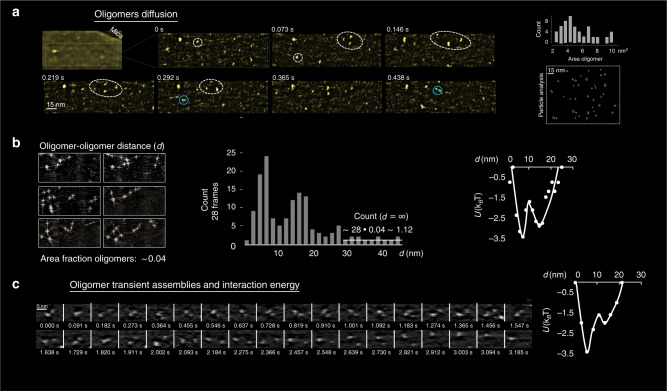
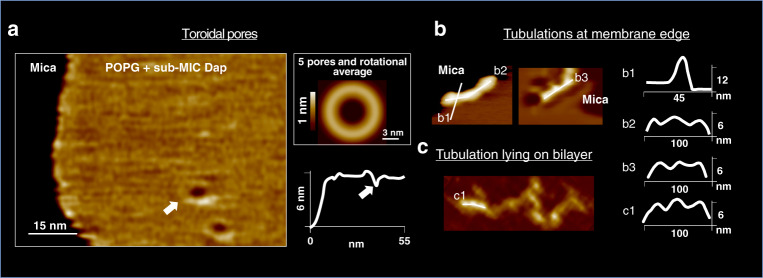
The increase in speed of the high-speed atomic force microscopy (HS-AFM) compared to that of the conventional AFM made possible the first-ever visualisation at the molecular-level of the activity of an antimicrobial peptide on a membrane. We investigated the medically prescribed but poorly understood lipopeptide Daptomycin under infection-like conditions (37 °C, bacterial lipid composition and antibiotic concentrations). We confirmed so far hypothetical models: Dap oligomerization and the existence of half pores. Moreover, we detected unknown molecular mechanisms: new mechanisms to form toroidal pores or to resist Dap action, and to unprecedently quantify the energy profile of interacting oligomers. Finally, the biological and medical relevance of the findings was ensured by a multi-scale multi-nativeness-from the molecule to the cell-correlation of molecular-level information from living bacteria (Bacillus subtilis strains) to liquid-suspended vesicles and supported-membranes using electron and optical microscopies and the lipid tension probe FliptR, where we found that the cells with a healthier state of their cell wall show smaller membrane deformations.
Conflict of interest statement
The authors declare no competing interests.
Figures






Similar articles
-
An Acyl-Linked Dimer of Daptomycin Is Strongly Inhibited by the Bacterial Cell Wall.ACS Infect Dis. 2017 Jul 14;3(7):462-466. doi: 10.1021/acsinfecdis.7b00019. Epub 2017 Mar 29. ACS Infect Dis. 2017. PMID: 28350438
-
Daptomycin-mediated reorganization of membrane architecture causes mislocalization of essential cell division proteins.J Bacteriol. 2012 Sep;194(17):4494-504. doi: 10.1128/JB.00011-12. Epub 2012 Jun 1. J Bacteriol. 2012. PMID: 22661688 Free PMC article.
-
Real-time multimodal imaging of daptomycin action on the cell wall of adherent Staphylococcus aureus.BMC Res Notes. 2025 Feb 5;18(1):54. doi: 10.1186/s13104-025-07130-5. BMC Res Notes. 2025. PMID: 39910624 Free PMC article.
-
Mechanisms of drug resistance: daptomycin resistance.Ann N Y Acad Sci. 2015 Sep;1354:32-53. doi: 10.1111/nyas.12948. Epub 2015 Oct 23. Ann N Y Acad Sci. 2015. PMID: 26495887 Free PMC article. Review.
-
[Molecular mechanisms underlying daptomycin resistance].Sheng Wu Gong Cheng Xue Bao. 2018 Jun 25;34(6):832-838. doi: 10.13345/j.cjb.170515. Sheng Wu Gong Cheng Xue Bao. 2018. PMID: 29943529 Review. Chinese.
Cited by
-
Metabolic engineering of Streptomyces roseosporus for increased production of clinically important antibiotic daptomycin.Microb Biotechnol. 2024 Nov;17(11):e70038. doi: 10.1111/1751-7915.70038. Microb Biotechnol. 2024. PMID: 39487765 Free PMC article.
-
Outer membrane and phospholipid composition of the target membrane affect the antimicrobial potential of first- and second-generation lipophosphonoxins.Sci Rep. 2021 May 17;11(1):10446. doi: 10.1038/s41598-021-89883-0. Sci Rep. 2021. PMID: 34001940 Free PMC article.
-
Visualizing the membrane disruption action of antimicrobial peptides by cryo-electron tomography.Nat Commun. 2023 Sep 6;14(1):5464. doi: 10.1038/s41467-023-41156-2. Nat Commun. 2023. PMID: 37673860 Free PMC article.
-
Recent Advances in Amphipathic Peptidomimetics as Antimicrobial Agents to Combat Drug Resistance.Molecules. 2024 May 24;29(11):2492. doi: 10.3390/molecules29112492. Molecules. 2024. PMID: 38893366 Free PMC article. Review.
-
Breeding of High Daptomycin-Producing Strain by Streptomycin Resistance Superposition.Pol J Microbiol. 2022 Sep 24;71(3):463-471. doi: 10.33073/pjm-2022-041. eCollection 2022 Sep 1. Pol J Microbiol. 2022. PMID: 36185027 Free PMC article.
References
-
- Reddy K, Yedery R, Aranha C. Antimicrobial peptides: premises and promises. Int. J. Antimicrob. Agents. 2004;24:536–547. - PubMed
-
- Boman H. Antibacterial peptides: basic facts and emerging concepts. J. Intern. Med. 2003;254:197–215. - PubMed
-
- Pirri G, Giuliani A, Nicoletto SF, Pizzuto L, Rinaldi AC. Lipopeptides as anti-infectives: a practical perspective. Cent. Eur. J. Biol. 2009;4:258–273.
-
- Hashizume, H. & Nishimura, Y. Studies in Natural Products Chemistry. Vol. 35, 693–751 (Elsevier, 2008).
-
- Yin N, et al. Structure–activity relationship studies of a series of semisynthetic lipopeptides leading to the discovery of Surotomycin, a novel cyclic lipopeptide being developed for the treatment of Clostridium difficile-associated diarrhea. J. Med. Chem. 2015;58:5137–5142. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
