

Wnt-inducible Lrp6-APEX2 interacting proteins identify ESCRT machinery and Trk-fused gene as components of the Wnt signaling pathway
- PMID: 33299006
- PMCID: PMC7726150
- DOI: 10.1038/s41598-020-78019-5
Wnt-inducible Lrp6-APEX2 interacting proteins identify ESCRT machinery and Trk-fused gene as components of the Wnt signaling pathway
Abstract
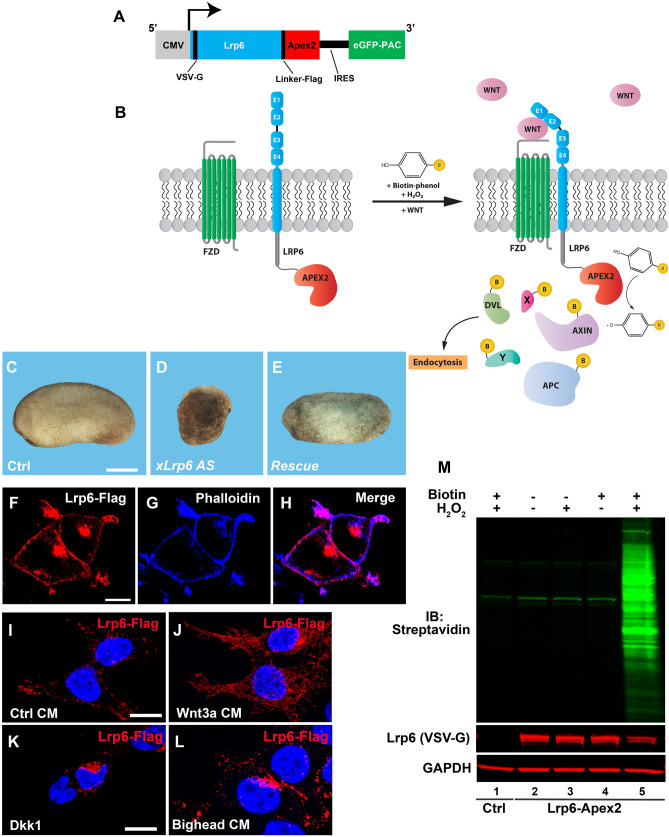
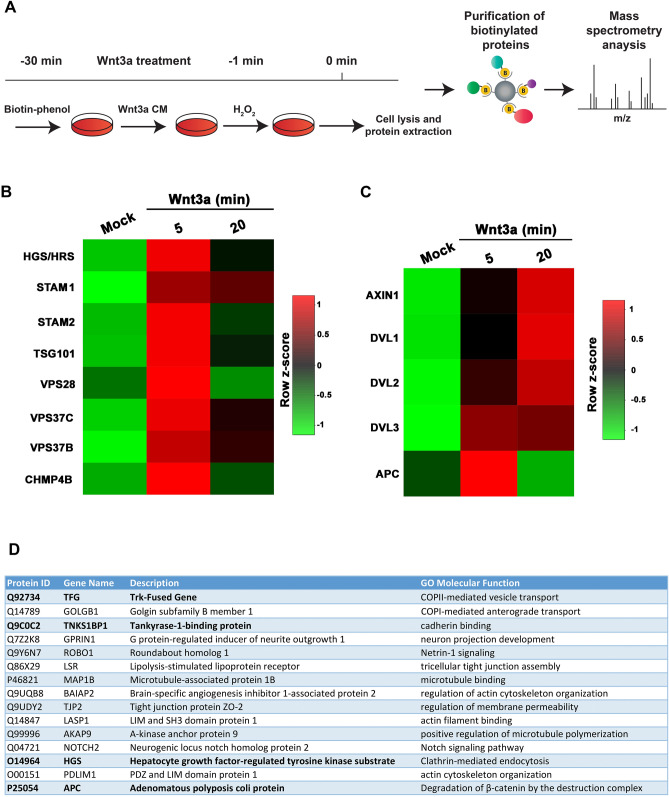
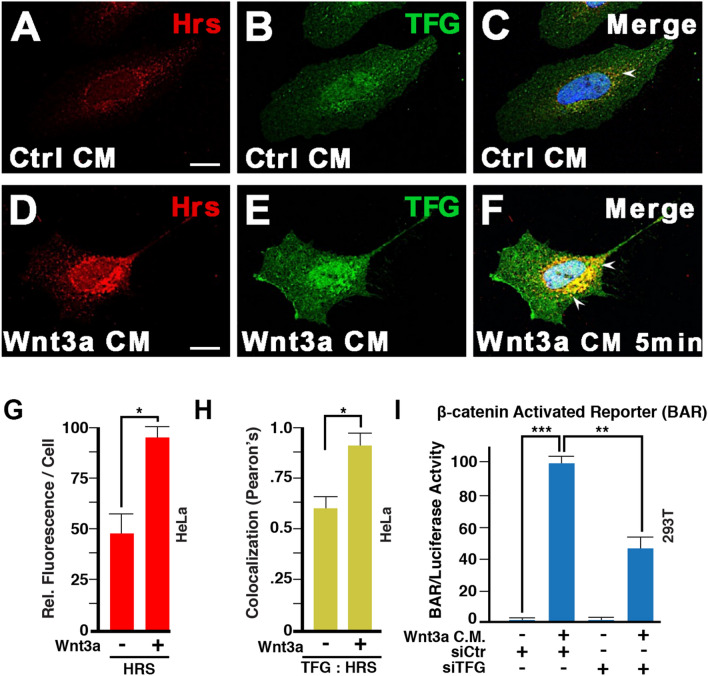
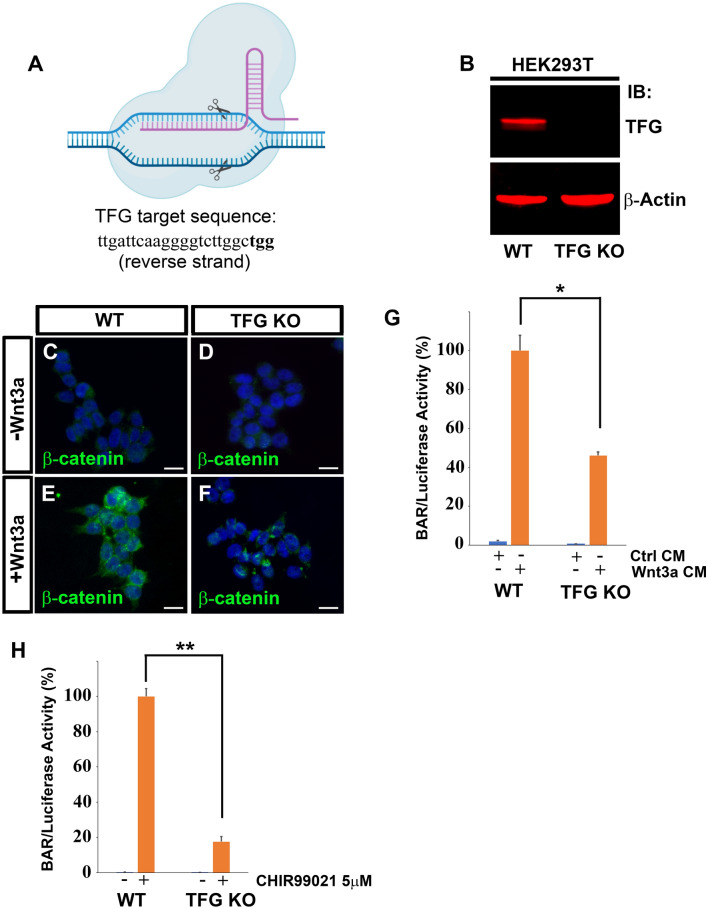
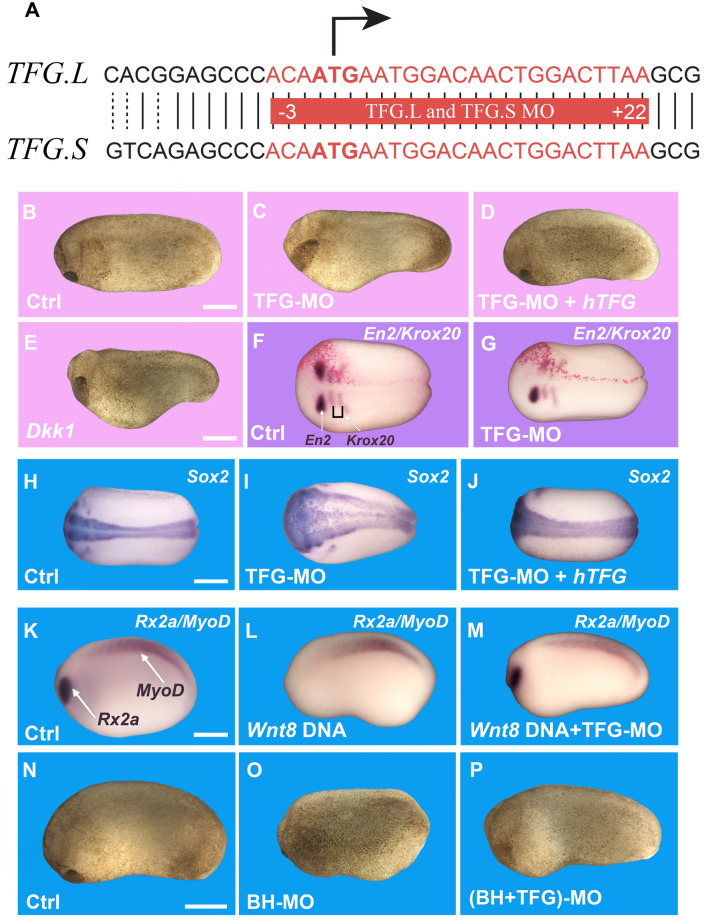
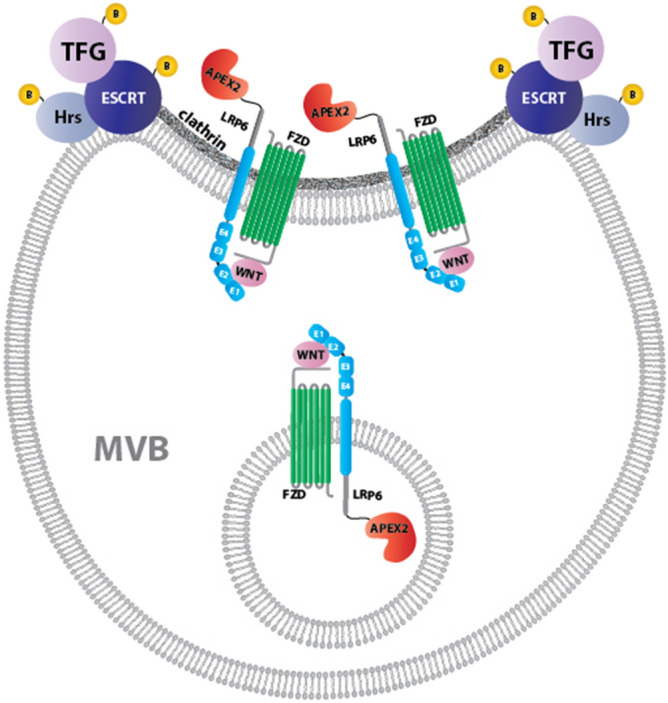
The canonical Wnt pathway serves as a hub connecting diverse cellular processes, including β-catenin signaling, differentiation, growth, protein stability, macropinocytosis, and nutrient acquisition in lysosomes. We have proposed that sequestration of β-catenin destruction complex components in multivesicular bodies (MVBs) is required for sustained canonical Wnt signaling. In this study, we investigated the events that follow activation of the canonical Wnt receptor Lrp6 using an APEX2-mediated proximity labeling approach. The Wnt co-receptor Lrp6 was fused to APEX2 and used to biotinylate targets that are recruited near the receptor during Wnt signaling at different time periods. Lrp6 proximity targets were identified by mass spectrometry, and revealed that many endosomal proteins interacted with Lrp6 within 5 min of Wnt3a treatment. Interestingly, we found that Trk-fused gene (TFG), previously known to regulate the cell secretory pathway and to be rearranged in thyroid and lung cancers, was strongly enriched in the proximity of Lrp6. TFG depletion with siRNA, or knock-out with CRISPR/Cas9, significantly reduced Wnt/β-catenin signaling in cell culture. In vivo, studies in the Xenopus system showed that TFG is required for endogenous Wnt-dependent embryonic patterning. The results suggest that the multivesicular endosomal machinery and the novel player TFG have important roles in Wnt signaling.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
