

Antigen-driven clonal selection shapes the persistence of HIV-1-infected CD4+ T cells in vivo
- PMID: 33301425
- PMCID: PMC7843227
- DOI: 10.1172/JCI145254
Antigen-driven clonal selection shapes the persistence of HIV-1-infected CD4+ T cells in vivo
Abstract
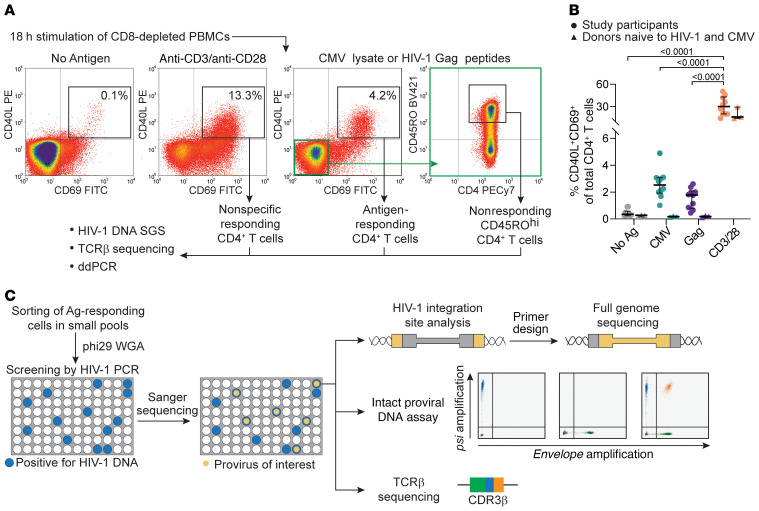
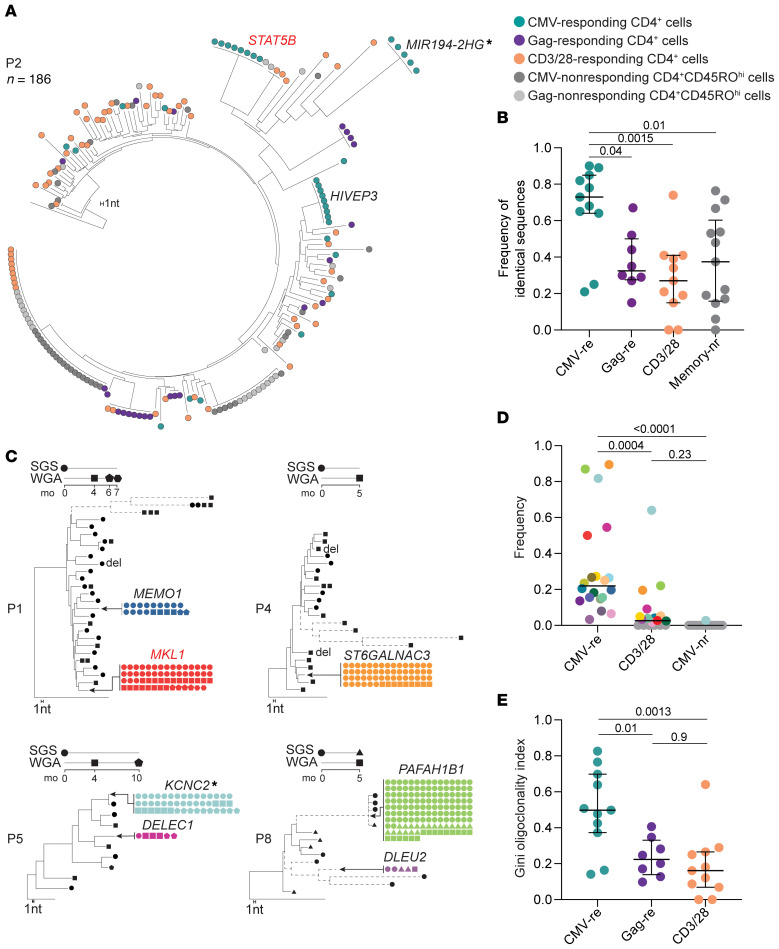
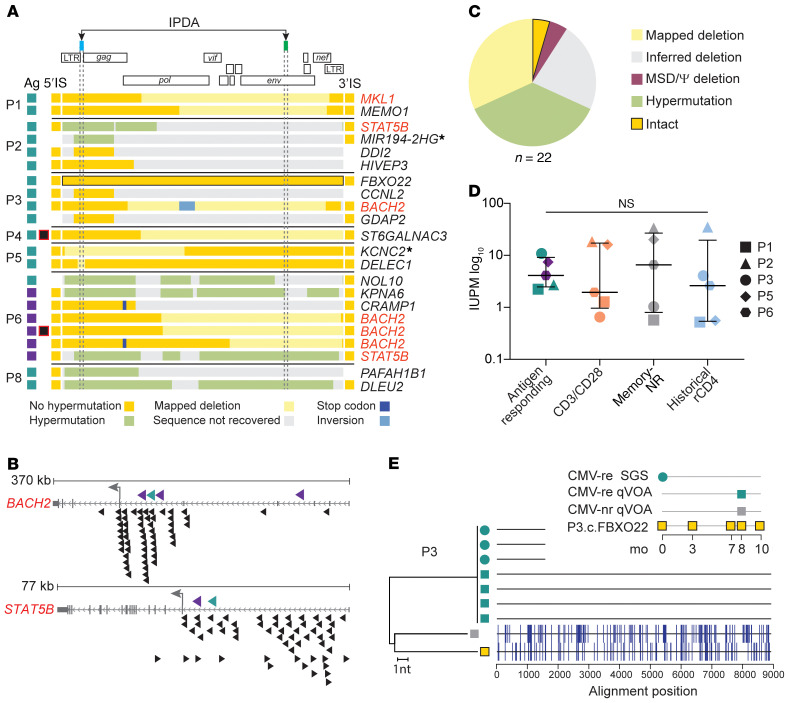
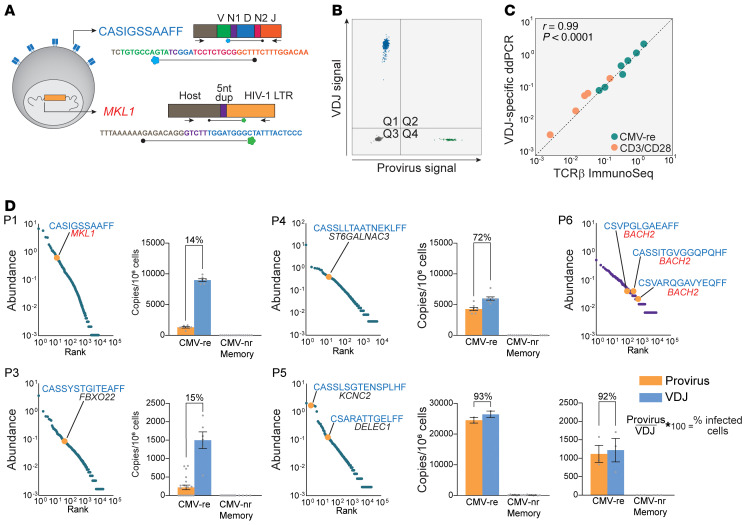
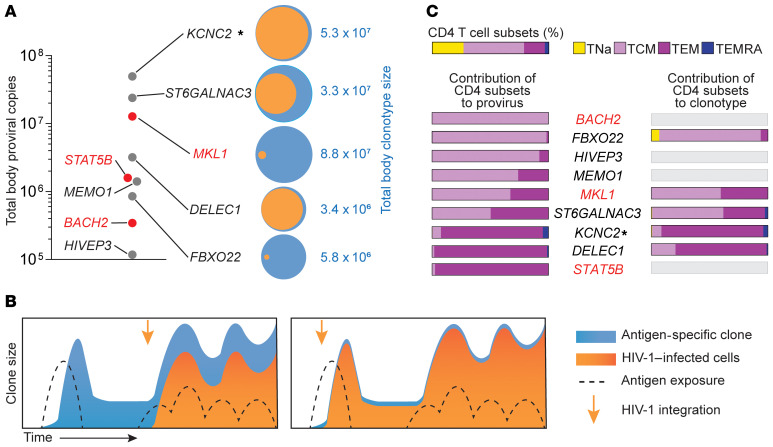
Clonal expansion of infected CD4+ T cells is a major mechanism of HIV-1 persistence and a barrier to achieving a cure. Potential causes are homeostatic proliferation, effects of HIV-1 integration, and interaction with antigens. Here, we show that it is possible to link antigen responsiveness, the full proviral sequence, the integration site, and the T cell receptor β-chain (TCRβ) sequence to examine the role of recurrent antigenic exposure in maintaining the HIV-1 reservoir. We isolated CMV- and Gag-responding CD4+ T cells from 10 treated individuals. Proviral populations in CMV-responding cells were dominated by large clones, including clones harboring replication-competent proviruses. TCRβ repertoires showed high clonality driven by converging adaptive responses. Although some proviruses were in genes linked to HIV-1 persistence (BACH2, STAT5B, MKL1), the proliferation of infected cells under antigenic stimulation occurred regardless of the site of integration. Paired TCRβ and integration site analysis showed that infection could occur early or late in the course of a clone's response to antigen and could generate infected cell populations too large to be explained solely by homeostatic proliferation. Together, these findings implicate antigen-driven clonal selection as a major factor in HIV-1 persistence, a finding that will be a difficult challenge to eradication efforts.
Keywords: AIDS/HIV; Adaptive immunity; Clonal selection; Immunology; T cells.
Conflict of interest statement
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
