

Phenotypic plasticity, genetic assimilation, and genetic compensation in hypoxia adaptation of high-altitude vertebrates
- PMID: 33301891
- PMCID: PMC7867641
- DOI: 10.1016/j.cbpa.2020.110865
Phenotypic plasticity, genetic assimilation, and genetic compensation in hypoxia adaptation of high-altitude vertebrates
Abstract
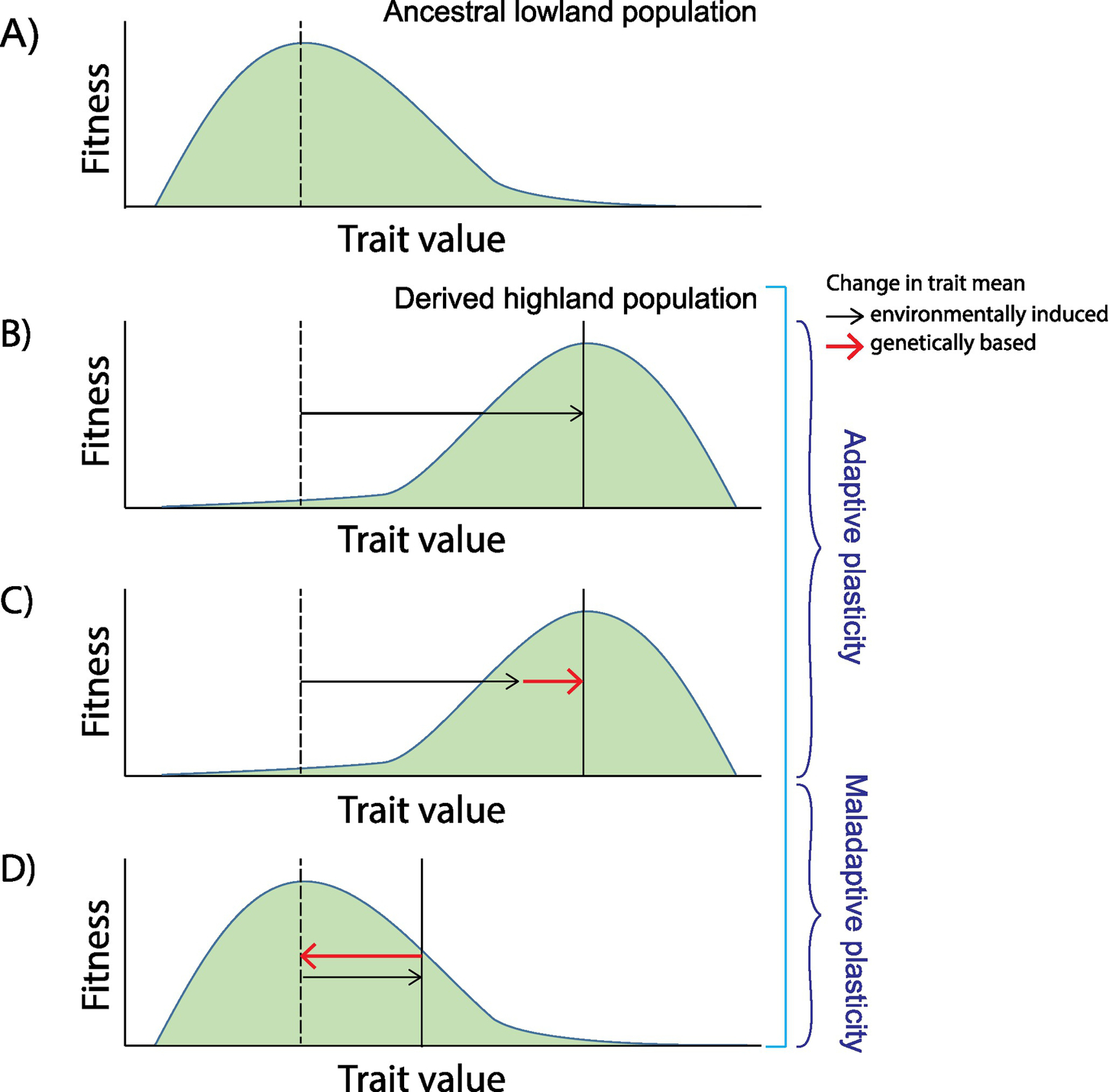
Important questions about mechanisms of physiological adaptation concern the role of phenotypic plasticity and the extent to which acclimatization responses align with genetic responses to selection. Such questions can be addressed in experimental studies of high-altitude vertebrates by investigating how mechanisms of acclimatization to hypoxia in lowland natives may influence genetic adaptation to hypoxia in highland natives. Evidence from high-altitude mammals suggest that evolved changes in some physiological traits involved canalization of the ancestral acclimatization response to hypoxia (genetic assimilation), a mechanism that results in an evolved reduction in plasticity. In addition to cases where adaptive plasticity may have facilitated genetic adaptation, evidence also suggests that some physiological changes in high-altitude natives are the result of selection to mitigate maladaptive plastic responses to hypoxia (genetic compensation). Examples of genetic compensation involve the attenuation of hypoxic pulmonary hypertension in Tibetan humans and other mammals native to high altitude. Here we discuss examples of adaptive physiological phenotypes in high-altitude natives that may have evolved by means of genetic assimilation or genetic compensation.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
We hereby state that we have no conflicts of interest.
Figures



References
-
- Brutsaert T, 2016. Why are high altitude natives so strong at high altitude? Nature vs. nurture: genetic factors vs. growth and development. Advances in Experimental Medicine and Biology 903, 101–112. - PubMed
-
- Brutsaert TD, 2008. Do high-altitude natives have enhanced exercise performance at altitude? Applied Physiology Nutrition and Metabolism 33, 582–592. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
