

Glucosylceramide Plays a Role in Fungal Germination, Lipid Raft Organization and Biofilm Adhesion of the Pathogenic Fungus Scedosporium aurantiacum
- PMID: 33302332
- PMCID: PMC7762401
- DOI: 10.3390/jof6040345
Glucosylceramide Plays a Role in Fungal Germination, Lipid Raft Organization and Biofilm Adhesion of the Pathogenic Fungus Scedosporium aurantiacum
Abstract
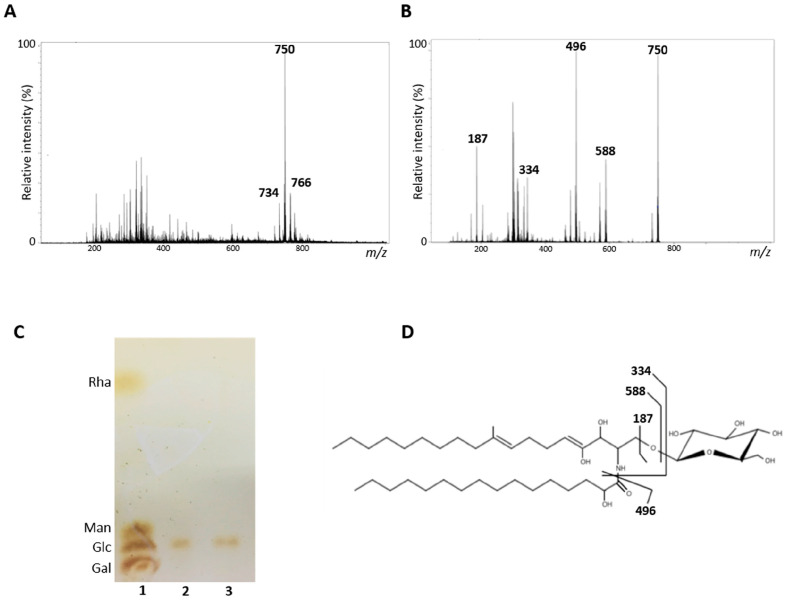
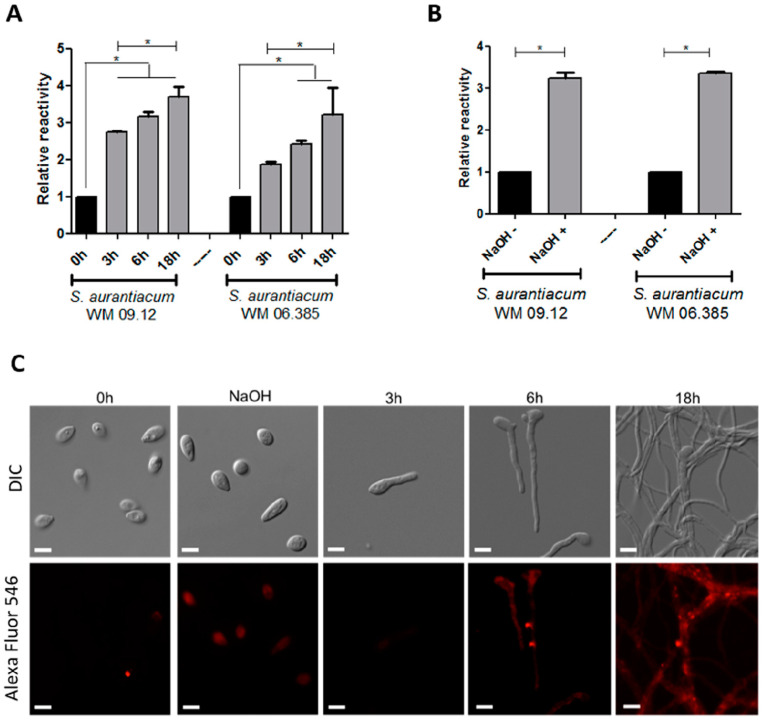
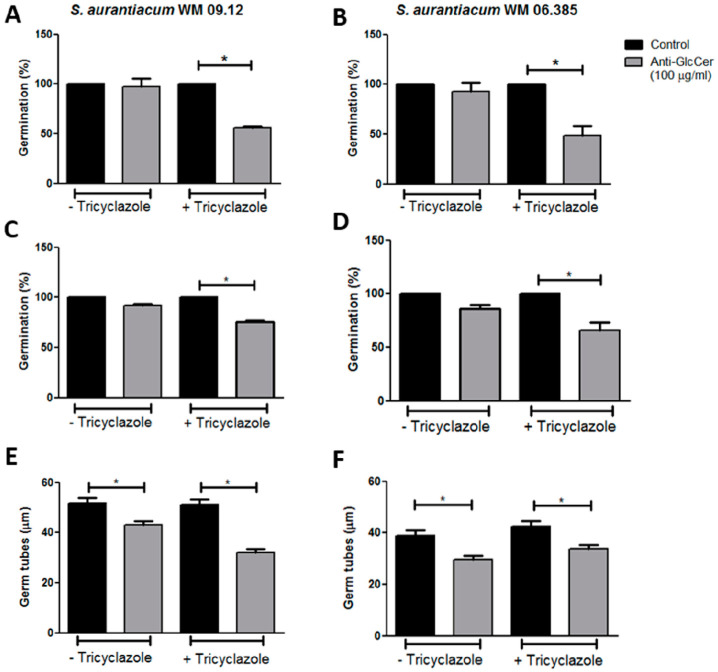
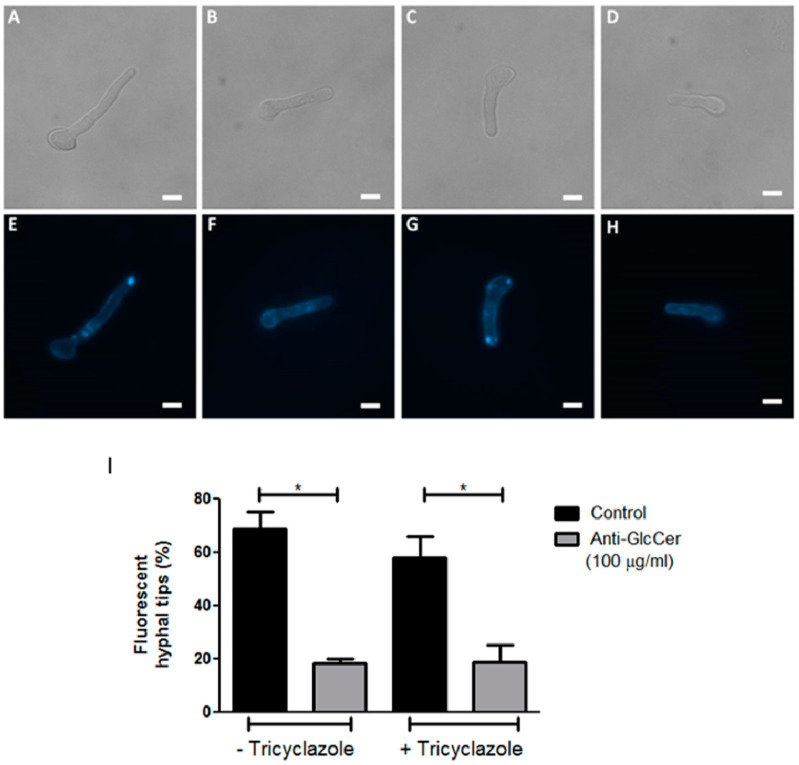
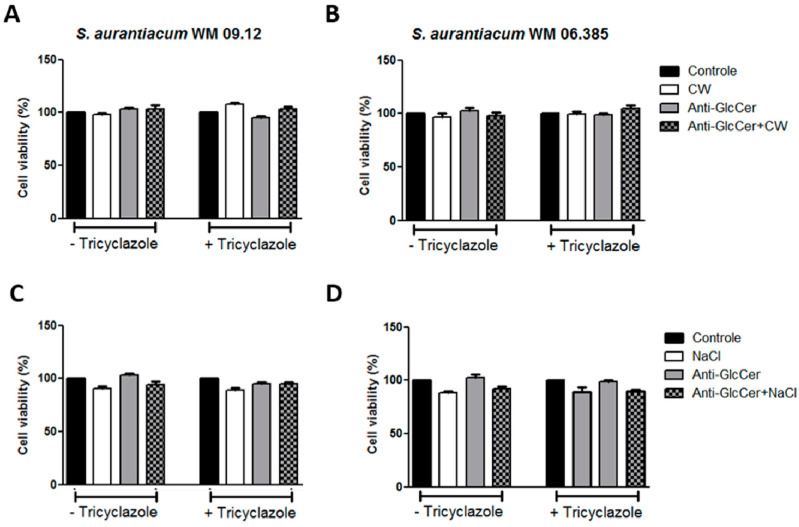
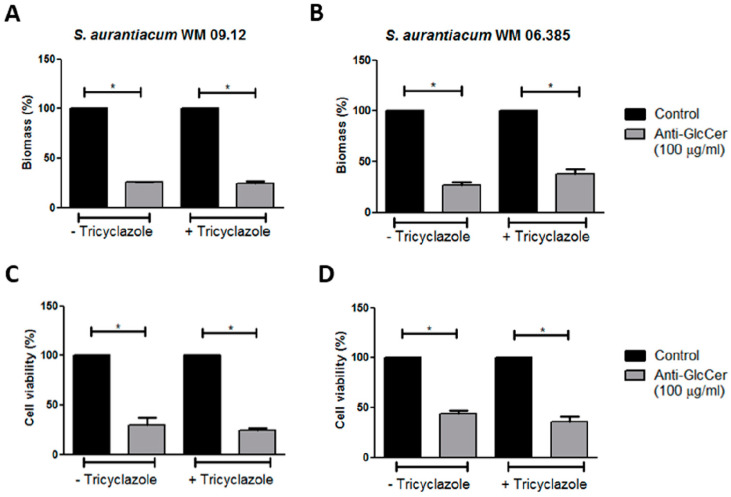
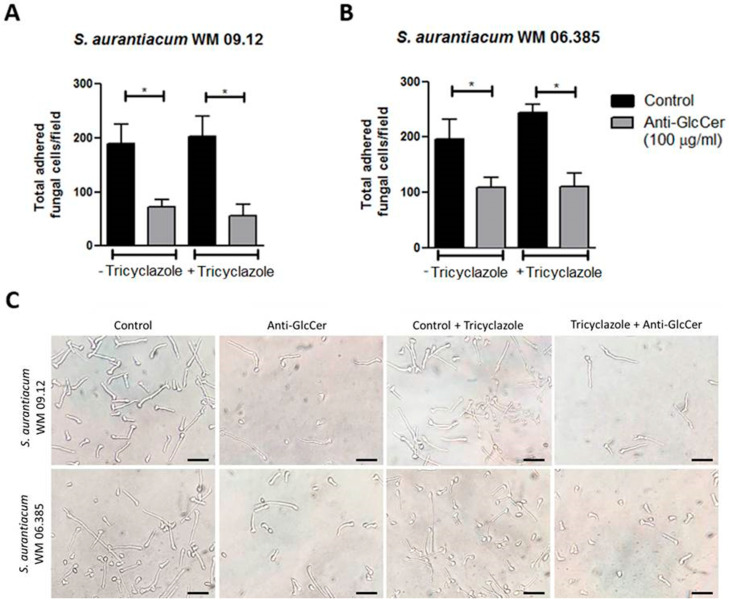
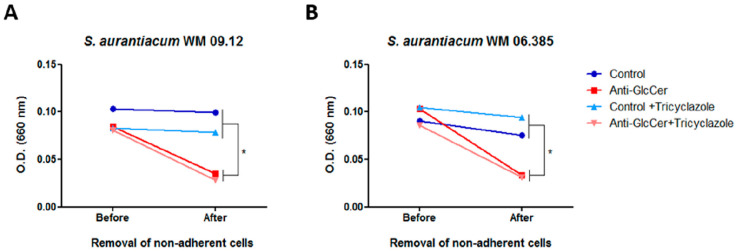
Infections caused by Scedosporium species present a wide range of clinical manifestations, from superficial to disseminated, especially in immunocompromised patients. Glucosylceramides (GlcCer) are glycosphingolipids found on the fungal cell surface and play an important role in growth and pathogenicity processes in different fungi. The present study aimed to evaluate the structure of GlcCer and its role during growth in two S. aurantiacum isolates. Purified GlcCer from both isolates were obtained and its chemical structure identified by mass spectrometry. Using ELISA and immunofluorescence techniques it was observed that germination and NaOH-treatment of conidia favor GlcCer exposure. Monoclonal anti-GlcCer antibody reduced germination when cultivated with the inhibitor of melanin synthesis tricyclazole and also reduced germ tube length of conidia, both cultivated or not with tricyclazole. It was also demonstrated that anti-GlcCer altered lipid rafts organization, as shown by using the fluorescent stain filipin, but did not affect the susceptibility of the cell surface to damaging agents. Anti-GlcCer reduced total biomass and viability in biofilms formed on polystyrene plates. In the presence of anti-GlcCer, germinated S. aurantiacum conidia and biofilms could not adhere to polystyrene with the same efficacy as control cells. These results highlight the relevance of GlcCer in growth processes of S. aurantiacum.
Keywords: Scedosporium; biofilm; fungal growth; glucosylceramide.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
LinkOut - more resources
Full Text Sources
