

Airway immunometabolites fuel Pseudomonas aeruginosa infection
- PMID: 33302964
- PMCID: PMC7731785
- DOI: 10.1186/s12931-020-01591-x
Airway immunometabolites fuel Pseudomonas aeruginosa infection
Abstract
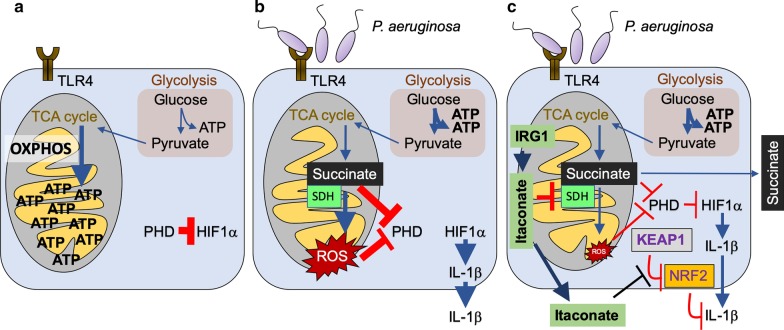
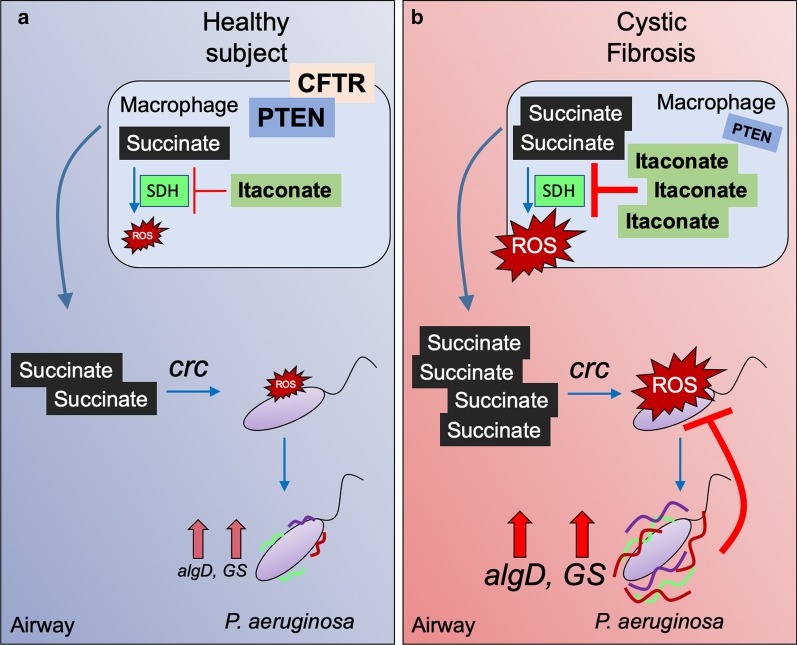
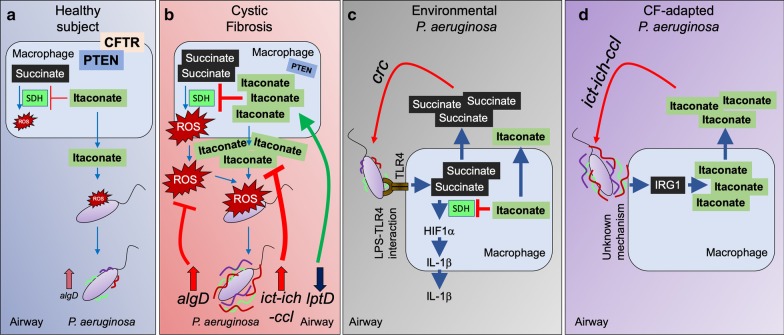
Pulmonary infections are associated with a brisk inflammatory reaction to bacterial surface components. Lipopolysaccharides (LPS) trigger macrophage activation and release of mitochondrial metabolites that control the intensity of the immune response. Whereas succinate induces oxidative stress (ROS), HIF1α stabilization, glycolysis and IL-1β release, itaconate suppresses inflammation by inhibiting succinate oxidation, glycolytic flux and promoting anti-oxidant Nrf2-HO-1 functions. P. aeruginosa is a major pathogen associated with acute and chronic lung infection. Although both secreted toxins, LPS and proteases are key factors to establish acute P. aeruginosa pneumonia, lack of these components in chronic P. aeruginosa isolates suggest these organisms exploit other mechanisms to adapt and persist in the lung. Upon inhalation, P. aeruginosa strains trigger airway macrophage reprograming and bacterial variants obtained from acutely and chronically infected subjects exhibit metabolic adaptation consistent with succinate and itaconate assimilation; namely, high expression of extracellular polysaccharides (EPS), reduced lptD-LPS function, increased glyoxylate shunt (GS) activity and substantial biofilm production. In this review we discuss recent findings illustrating how P. aeruginosa induces and adapts to macrophage metabolites in the human lung, and that catabolism of succinate and itaconate contribute to their formidable abilities to tolerate oxidative stress, phagocytosis and immune clearance.
Keywords: Adaptation; Biofilm; Cystic fibrosis; Immunometabolism; Itaconate; Metabolic stress; Pneumonia; Pseudomonas aeruginosa; ROS; Succinate.
Conflict of interest statement
Authors declare no conflicts of interest exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
