

Somatostatin expressing GABAergic interneurons in the medial entorhinal cortex preferentially inhibit layerIII-V pyramidal cells
- PMID: 33303963
- PMCID: PMC7728756
- DOI: 10.1038/s42003-020-01496-x
Somatostatin expressing GABAergic interneurons in the medial entorhinal cortex preferentially inhibit layerIII-V pyramidal cells
Abstract
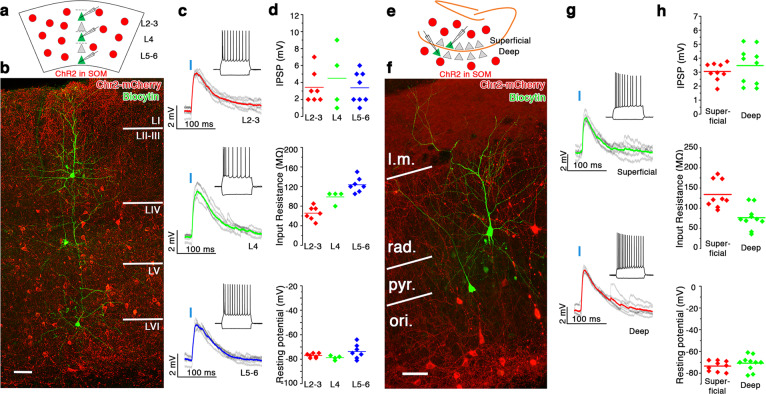
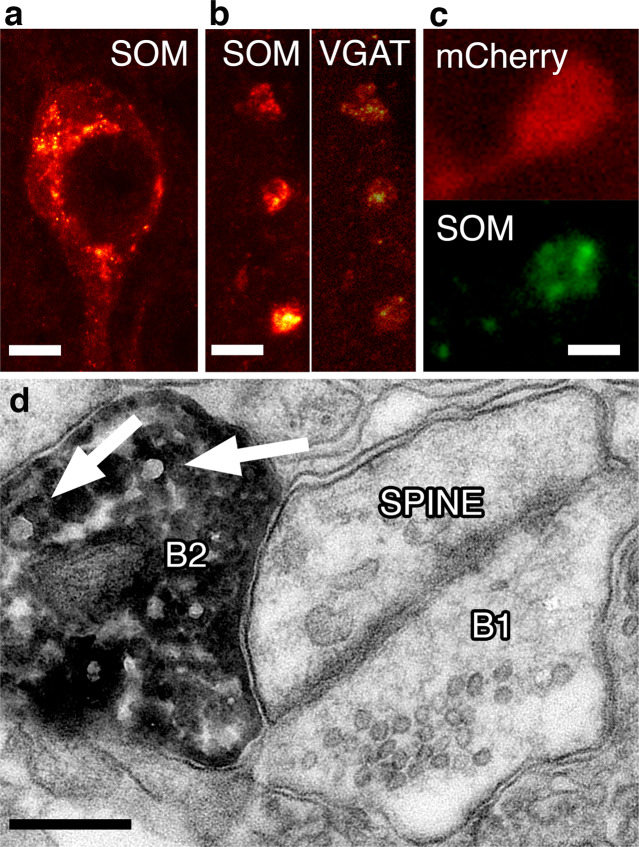
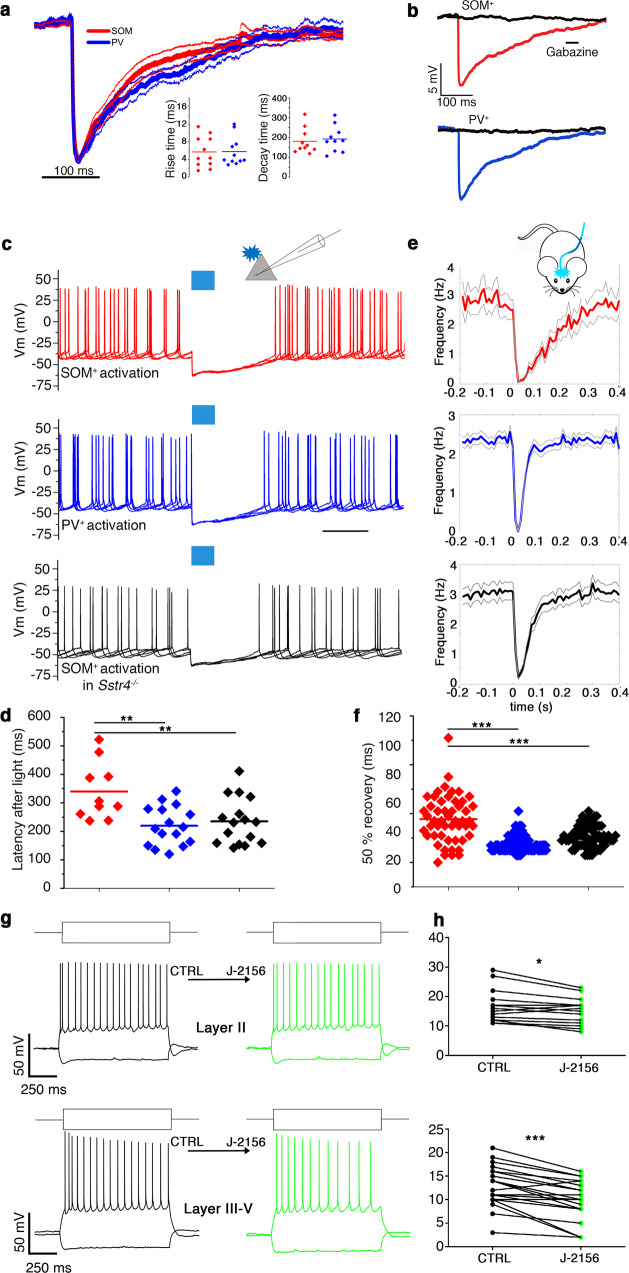
GABA released from heterogeneous types of interneurons acts in a complex spatio-temporal manner on postsynaptic targets in the networks. In addition to GABA, a large fraction of GABAergic cells also express neuromodulator peptides. Somatostatin (SOM) containing interneurons, in particular, have been recognized as key players in several brain circuits, however, the action of SOM and its downstream network effects remain largely unknown. Here, we used optogenetics, electrophysiologic, anatomical and behavioral experiments to reveal that the dendrite-targeting, SOM+ GABAergic interneurons demonstrate a unique layer-specific action in the medial entorhinal cortex (MEC) both in terms of GABAergic and SOM-related properties. We show that GABAergic and somatostatinergic neurotransmission originating from SOM+ local interneurons preferentially inhibit layerIII-V pyramidal cells, known to be involved in memory formation. We propose that this dendritic GABA-SOM dual inhibitory network motif within the MEC serves to selectively modulate working-memory formation without affecting the retrieval of already learned spatial navigation tasks.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
