

Functions and regulation of the serine/threonine protein kinase CK1 family: moving beyond promiscuity
- PMID: 33306089
- PMCID: PMC7733671
- DOI: 10.1042/BCJ20200506
Functions and regulation of the serine/threonine protein kinase CK1 family: moving beyond promiscuity
Abstract
Regarded as constitutively active enzymes, known to participate in many, diverse biological processes, the intracellular regulation bestowed on the CK1 family of serine/threonine protein kinases is critically important, yet poorly understood. Here, we provide an overview of the known CK1-dependent cellular functions and review the emerging roles of CK1-regulating proteins in these processes. We go on to discuss the advances, limitations and pitfalls that CK1 researchers encounter when attempting to define relationships between CK1 isoforms and their substrates, and the challenges associated with ascertaining the correct physiological CK1 isoform for the substrate of interest. With increasing interest in CK1 isoforms as therapeutic targets, methods of selectively inhibiting CK1 isoform-specific processes is warranted, yet challenging to achieve given their participation in such a vast plethora of signalling pathways. Here, we discuss how one might shut down CK1-specific processes, without impacting other aspects of CK1 biology.
Keywords: FAM83; Wnt proteins; casein kinase; cell cycle; circadian clock.
© 2020 The Author(s).
Conflict of interest statement
The authors declare that there are no competing interests associated with the manuscript.
Figures
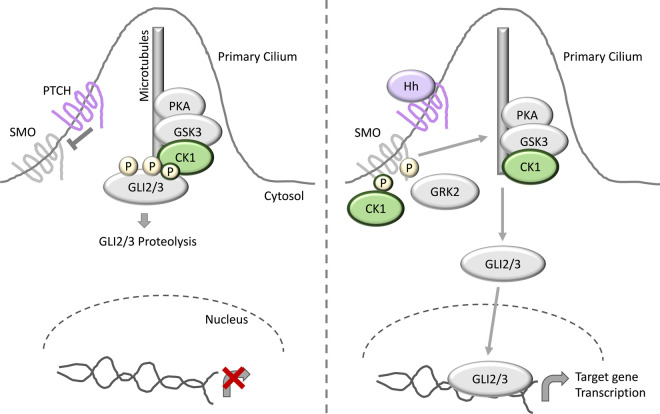
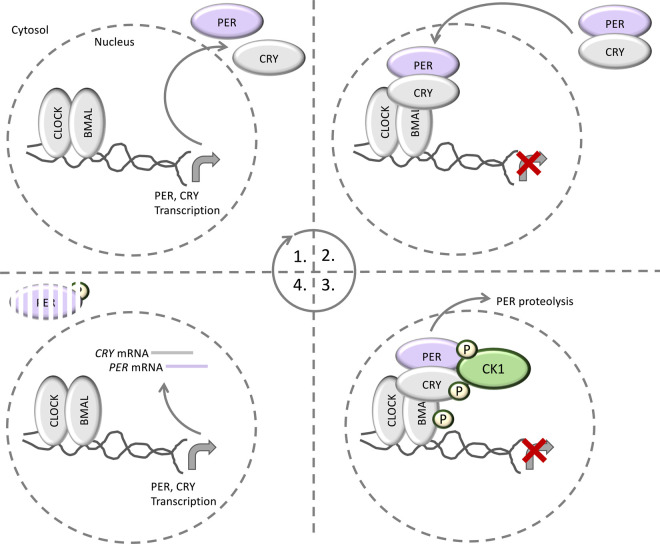
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials